

doi: 10.1128/JB.187.4.1287-1292.2005.
A polysaccharide deacetylase homologue, PdaA, in Bacillus subtilis acts as an N-acetylmuramic acid deacetylase in vitro
Affiliations
- PMID: 15687192
- PMCID: PMC545626
- DOI: 10.1128/JB.187.4.1287-1292.2005
Item in Clipboard
A polysaccharide deacetylase homologue, PdaA, in Bacillus subtilis acts as an N-acetylmuramic acid deacetylase in vitro
J Bacteriol.
2005 Feb.
Abstract
A polysaccharide deacetylase homologue, PdaA, was determined to act as an N-acetylmuramic acid deacetylase in vitro. Histidine-tagged truncated PdaA (with the putative signal sequence removed) was overexpressed in Escherichia coli cells and purified. Measurement of deacetylase activity showed that PdaA could deacetylate peptidoglycan treated with N-acetylmuramoyl-L-alanine amidase CwlH but could not deacetylate peptidoglycan treated with or without DL-endopeptidase LytF (CwlE). Reverse-phase high-performance liquid chromatography and mass spectrometry (MS) and MS-MS analyses indicated that PdaA could deacetylate the N-acetylmuramic acid residues of purified glycan strands derived from Bacillus subtilis peptidoglycan.
Figures
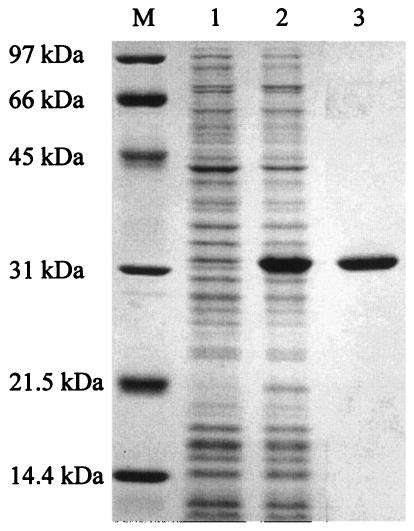
SDS-14% PAGE of h-ΔPdaA. h-ΔPdaA was expressed in E. coli JM109 cells in the presence of 1 mM IPTG. Lanes 1 and 2, proteins extracted from whole cells without and with IPTG added, respectively; lane 3, purified h-ΔPdaA; lane M, protein standards (Bio-Rad).
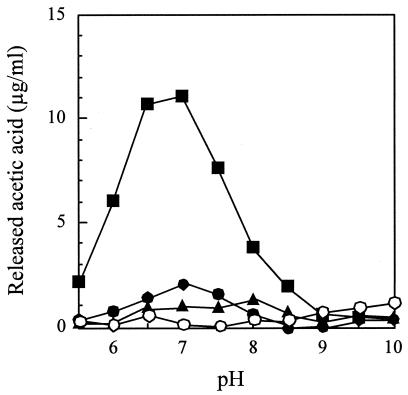
Deacetylase activity of h-ΔPdaA with nontreated peptidoglycan (•), peptidoglycan treated with l -alanine amidase CwlH (▪), and peptidoglycan treated with dl -endopeptidase LytF (▴). One milligram of chitin oligomer (hexa-N-acetylchitohexaose) per ml was also used (○). Purified h-ΔPdaA (final concentration, 2 μg/ml) was added to the following buffers (final concentration, 50 mM) containing 1 mg of nontreated peptidoglycan per ml or peptidoglycan treated with CwlH or LytF as a substrate (see Materials and Methods): morpholineethanesulfonic acid (MES) buffer for pH 5.5, 6.0, and 6.5; HEPES buffer for pH 7.0, 7.5, 8.0, and 8.5; and CHES [2-(cyclohexylamino)-ethanesulfonic acid] buffer for pH 9.0, 9.5, and 10.0. The deacetylase reaction was carried out at 37°C for 4 h.
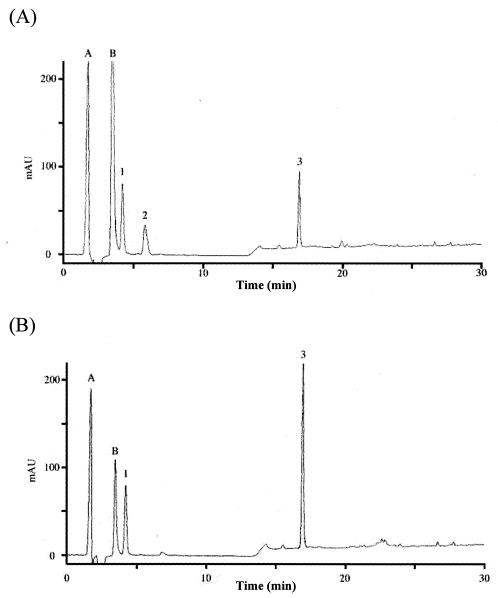
Lysozyme digestion of isolated N-acetylated peptidoglycan glycan strands treated with (A) or without (B) PdaA. Peptidoglycan from B. subtilis 168 was digested with l -alanine amidase CwlH, and then N-acetylated glycan strands were purified mainly by size exclusion chromatography (see Materials and Methods). After the N-acetylated glycan strands had been treated with (A) or without (B) PdaA, samples were treated with lysozyme, and this was followed by addition of NaBH4. The samples were separated by RP-HPLC. Peak 1, disaccharide (GlcNAc-MurNAcr); peak 2, N-deacetylated tetrasaccharide (GlcNAc-Mur-GlcNAc-MurNAcr); peak 3, tetrasaccharide (GlcNAc-MurNAc-GlcNAc-MurNAcr); peak A, reaction buffer; peak B, NaN3.
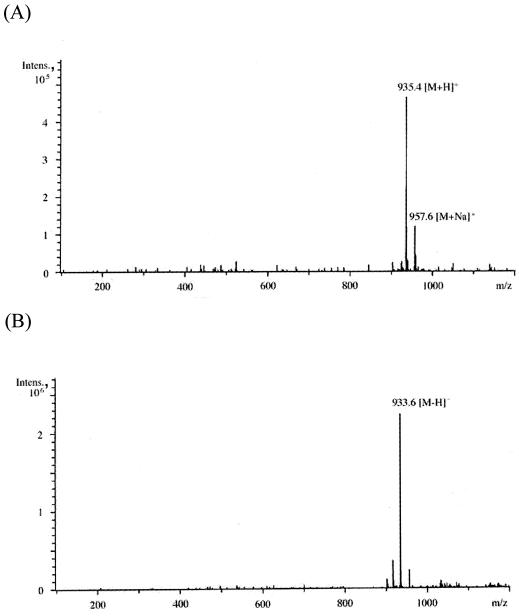
(A to E) ESI-MS analyses (A and B) and ESI-MS-MS analyses (C to E) of peak 2 (GlcNAc-Mur-GlcNAc-MurNAcr) in Fig. 3A. Panels A and B show ESI-MS analysis in the positive and negative modes, respectively. Panels C to E show ESI-MS-MS analyses of the [M+H]+ precursor ion (m/z 935.4), [M+Na]+ precursor ion (m/z 957.6), and [M-H]− precursor ion (m/z 933.6), respectively. (F) Putative structure and calculated molecular weight of each fragment of peak 2 in Fig. 3A. Ion series b, y, a, and z correspond to the fragment peaks in panels C to E. Intens., intensity.
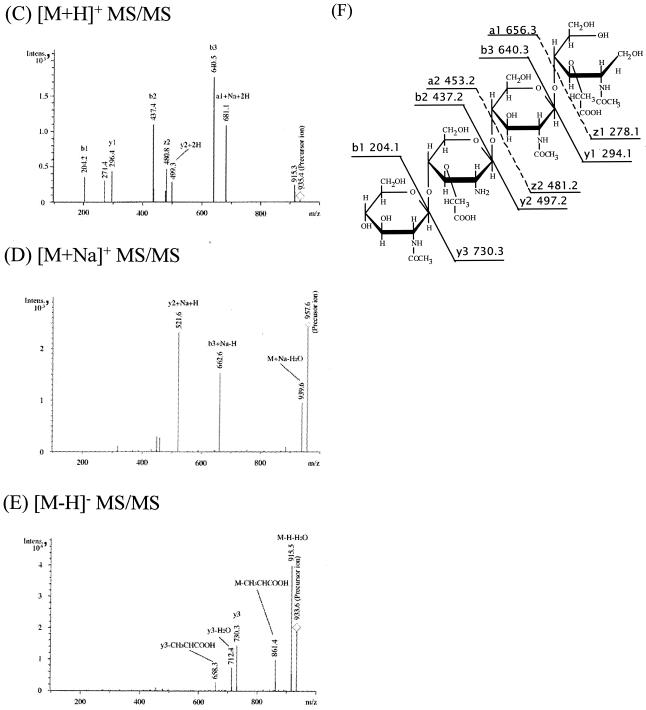
(A to E) ESI-MS analyses (A and B) and ESI-MS-MS analyses (C to E) of peak 2 (GlcNAc-Mur-GlcNAc-MurNAcr) in Fig. 3A. Panels A and B show ESI-MS analysis in the positive and negative modes, respectively. Panels C to E show ESI-MS-MS analyses of the [M+H]+ precursor ion (m/z 935.4), [M+Na]+ precursor ion (m/z 957.6), and [M-H]− precursor ion (m/z 933.6), respectively. (F) Putative structure and calculated molecular weight of each fragment of peak 2 in Fig. 3A. Ion series b, y, a, and z correspond to the fragment peaks in panels C to E. Intens., intensity.
References
-
- Blair, D. E., and D. M. van Aalten. 2004. Structures of Bacillus subtilis PdaA, a family 4 carbohydrate esterase, and a complex with N-acetyl-glucosamine. FEBS Lett. 570:13-19. - PubMed
-
- Fukushima, T., S. Ishikawa, H. Yamamoto, N. Ogasawara, and J. Sekiguchi. 2003. Transcriptional, functional and cytochemical analyses of the veg gene in Bacillus subtilis. J. Biochem. 133:475-483. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
