

Signalling crosstalk in FGF2-mediated protection of endothelial cells from HIV-gp120
- PMID: 15689238
- PMCID: PMC549045
- DOI: 10.1186/1471-2202-6-8
Signalling crosstalk in FGF2-mediated protection of endothelial cells from HIV-gp120
Abstract
Background: The blood brain barrier (BBB) is the first line of defence of the central nervous system (CNS) against circulating pathogens, such as HIV. The cytotoxic HIV protein, gp120, damages endothelial cells of the BBB, thereby compromising its integrity, which may lead to migration of HIV-infected cells into the brain. Fibroblast growth factor 2 (FGF2), produced primarily by astrocytes, promotes endothelial cell fitness and angiogenesis. We hypothesized that treatment of human umbilical vein endothelial cells (HUVEC) with FGF2 would protect the cells from gp120-mediated toxicity via endothelial cell survival signalling.
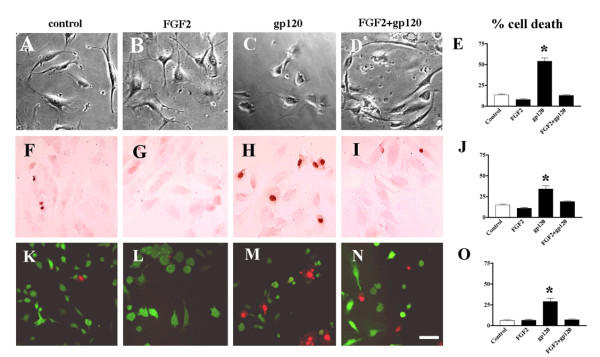
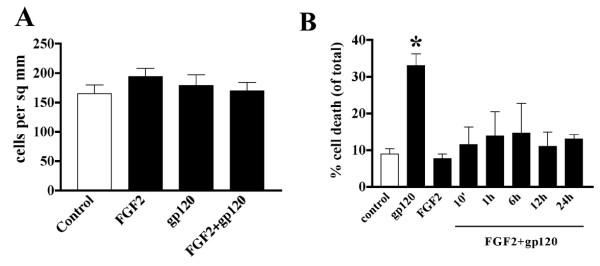
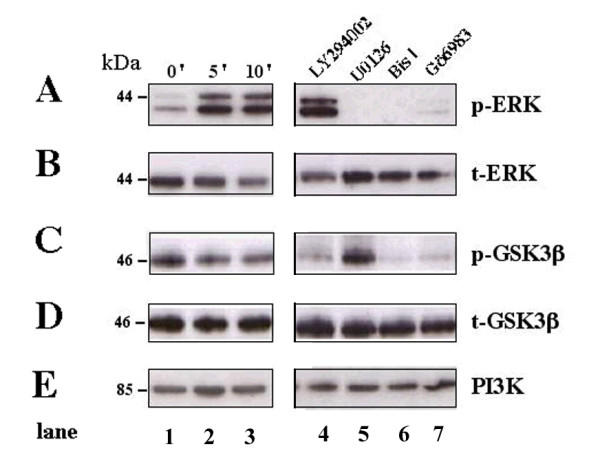
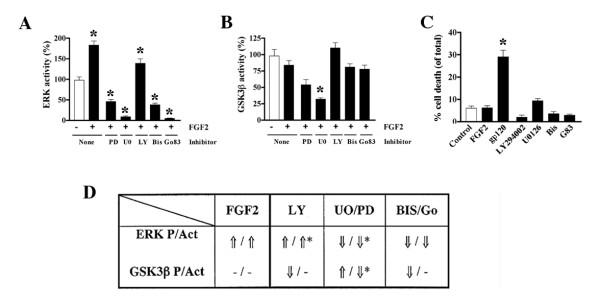
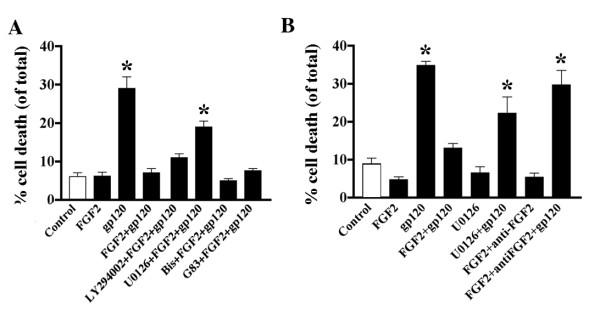
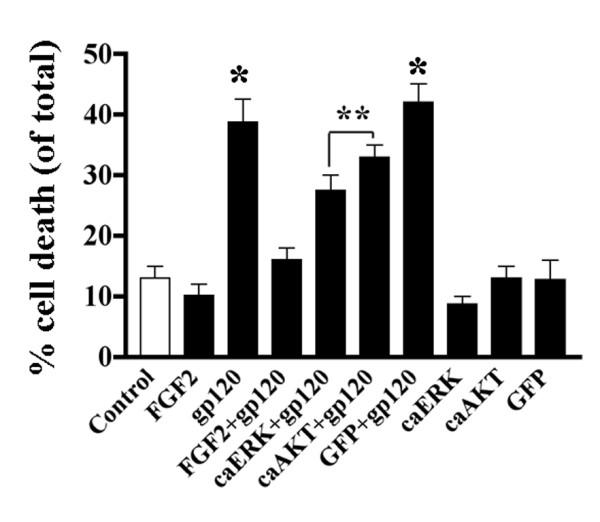
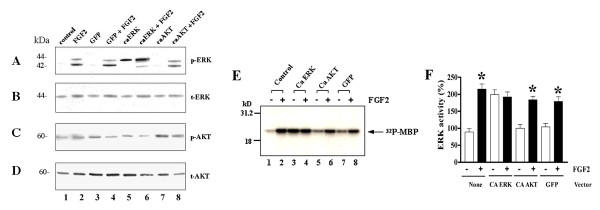
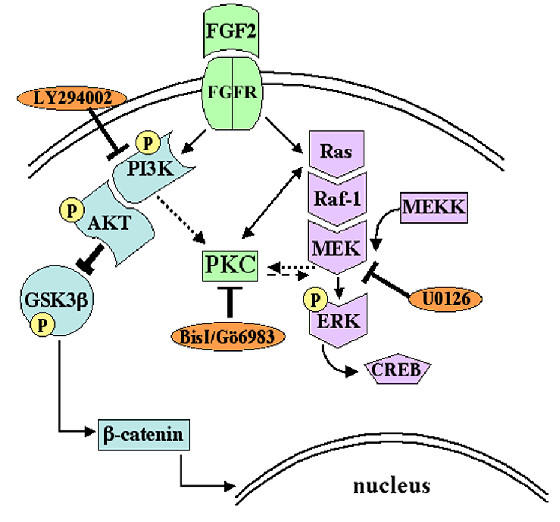
Results: Exposure of HUVEC to gp120 resulted in dose- and time-dependent cell death; whereas, pre-treatment of endothelial cells with FGF2 protected cells from gp120 angiotoxicity. Treatment of HUVEC with FGF2 resulted in dose- and time-dependent activation of the extracellular regulated kinase (ERK), with moderate effects on phosphoinositol 3 kinase (PI3K) and protein kinase B (PKB), also known as AKT, but no effects on glycogen synthase kinase 3 (GSK3beta) activity. Using pharmacological approaches, gene transfer and kinase activity assays, we show that FGF2-mediated angioprotection against gp120 toxicity is regulated by crosstalk among the ERK, PI3K-AKT and PKC signalling pathways.
Conclusions: Taken together, these results suggest that FGF2 may play a significant role in maintaining the integrity of the BBB during the progress of HIV associated cerebral endothelial cell damage.
Figures

Similar articles
-
HIV-1 gp120 compromises blood-brain barrier integrity and enhances monocyte migration across blood-brain barrier: implication for viral neuropathogenesis.J Cereb Blood Flow Metab. 2007 Jan;27(1):123-34. doi: 10.1038/sj.jcbfm.9600330. Epub 2006 May 10. J Cereb Blood Flow Metab. 2007. PMID: 16685256 Free PMC article.
-
Dysregulation of signal transduction pathways as a potential mechanism of nervous system alterations in HIV-1 gp120 transgenic mice and humans with HIV-1 encephalitis.J Clin Invest. 1996 Feb 1;97(3):789-98. doi: 10.1172/JCI118478. J Clin Invest. 1996. PMID: 8609236 Free PMC article.
-
HIV-1 gp120 induces cytokine expression, leukocyte adhesion, and transmigration across the blood-brain barrier: modulatory effects of STAT1 signaling.Microvasc Res. 2009 Mar;77(2):212-9. doi: 10.1016/j.mvr.2008.11.003. Epub 2008 Nov 28. Microvasc Res. 2009. PMID: 19103208 Free PMC article.
-
Blood-brain barrier abnormalities caused by HIV-1 gp120: mechanistic and therapeutic implications.ScientificWorldJournal. 2012;2012:482575. doi: 10.1100/2012/482575. Epub 2012 Feb 1. ScientificWorldJournal. 2012. PMID: 22448134 Free PMC article. Review.
-
Mechanisms of Blood-Retinal Barrier Disruption by HIV-1.Curr HIV Res. 2019;17(1):26-32. doi: 10.2174/1570162X17666190315163514. Curr HIV Res. 2019. PMID: 30873925 Review.
Cited by
-
Abbreviated Goal Management Training Shows Preliminary Evidence as a Neurorehabilitation Tool for HIV-associated Neurocognitive Disorders among Substance Users.Clin Neuropsychol. 2016;30(1):107-30. doi: 10.1080/13854046.2015.1129437. Epub 2016 Jan 12. Clin Neuropsychol. 2016. PMID: 26753986 Free PMC article. Clinical Trial.
-
Morphine treatment of human monocyte-derived macrophages induces differential miRNA and protein expression: impact on inflammation and oxidative stress in the central nervous system.J Cell Biochem. 2010 Jul 1;110(4):834-45. doi: 10.1002/jcb.22592. J Cell Biochem. 2010. PMID: 20564181 Free PMC article.
-
PTEN gene silencing prevents HIV-1 gp120(IIIB)-induced degeneration of striatal neurons.J Neurovirol. 2011 Feb;17(1):41-9. doi: 10.1007/s13365-010-0016-z. Epub 2011 Jan 14. J Neurovirol. 2011. PMID: 21234828 Free PMC article.
-
Zika virus induces FOXG1 nuclear displacement and downregulation in human neural progenitors.Stem Cell Reports. 2022 Jul 12;17(7):1683-1698. doi: 10.1016/j.stemcr.2022.05.008. Epub 2022 Jun 16. Stem Cell Reports. 2022. PMID: 35714598 Free PMC article.
-
Neuronal injury in simian immunodeficiency virus and other animal models of neuroAIDS.J Neurovirol. 2008 Aug;14(4):327-39. doi: 10.1080/13550280802132840. J Neurovirol. 2008. PMID: 18780234 Free PMC article. Review.
References
-
- Resnick L, Berger JR, Shapshak P, Tourtellotte WW. Early penetration of the blood-brain-barrier by HIV. Neurology. 1988;38:9–14. - PubMed
-
- Morris L, Silber E, Sonnenberg P, Eintracht S, Nyoka S, Lyons SF, Saffer D, Koornhof H, Martin DJ. High human immunodeficiency virus type 1 RNA load in the cerebrospinal fluid from patients with lymphocytic meningitis. J Infect Dis. 1998;177:473–476. - PubMed
-
- Budka H, Wiley CA, Kleihues P, Artigas J, Ashbury AK, Cho ES, Cornblath DR. HIV-associated disease of the nervous system: Review of nomenclature and proposal for neuropathology-based terminology. Brain Pathol. 1991;1:143–152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
