

The pleckstrin homology domain proteins Slm1 and Slm2 are required for actin cytoskeleton organization in yeast and bind phosphatidylinositol-4,5-bisphosphate and TORC2
- PMID: 15689497
- PMCID: PMC1073669
- DOI: 10.1091/mbc.e04-07-0564
The pleckstrin homology domain proteins Slm1 and Slm2 are required for actin cytoskeleton organization in yeast and bind phosphatidylinositol-4,5-bisphosphate and TORC2
Abstract
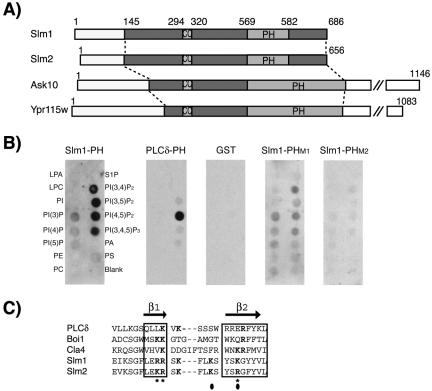
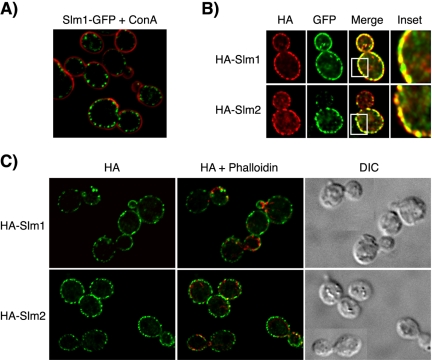
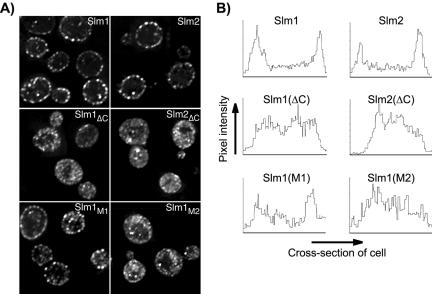
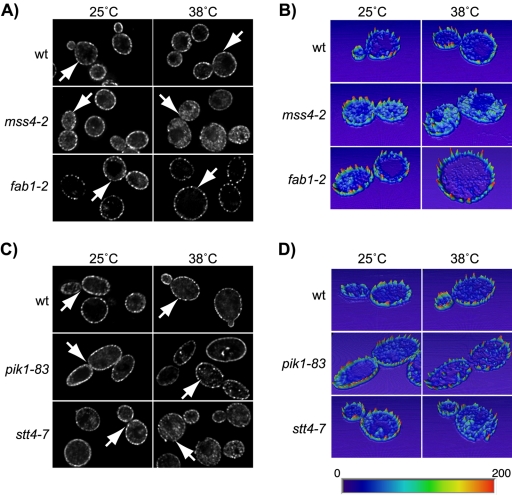
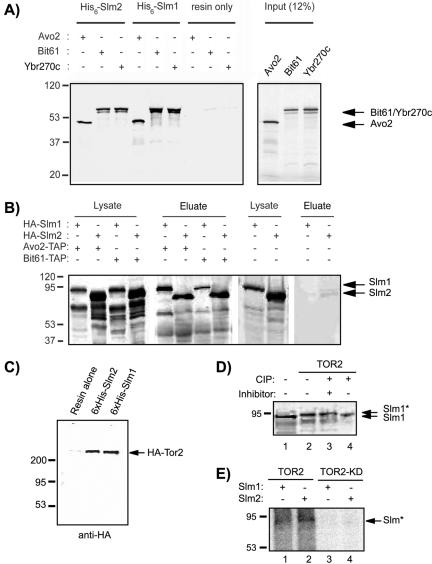
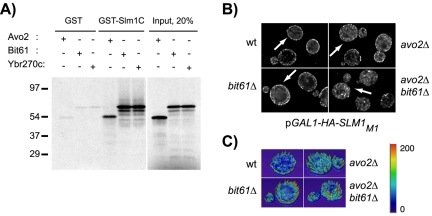
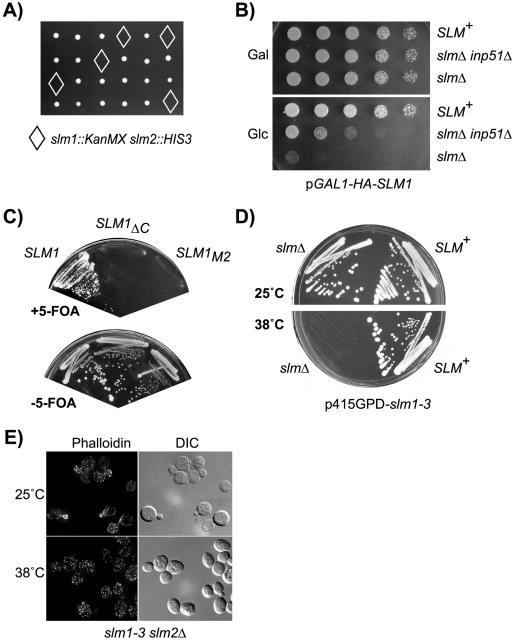
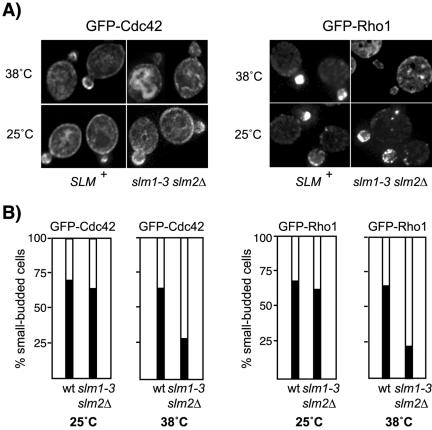
Phosphatidylinositol-4,5-bisphosphate [PtdIns(4,5)P(2)] is a key second messenger that regulates actin and membrane dynamics, as well as other cellular processes. Many of the effects of PtdIns(4,5)P(2) are mediated by binding to effector proteins that contain a pleckstrin homology (PH) domain. Here, we identify two novel effectors of PtdIns(4,5)P(2) in the budding yeast Saccharomyces cerevisiae: the PH domain containing protein Slm1 and its homolog Slm2. Slm1 and Slm2 serve redundant roles essential for cell growth and actin cytoskeleton polarization. Slm1 and Slm2 bind PtdIns(4,5)P(2) through their PH domains. In addition, Slm1 and Slm2 physically interact with Avo2 and Bit61, two components of the TORC2 signaling complex, which mediates Tor2 signaling to the actin cytoskeleton. Together, these interactions coordinately regulate Slm1 targeting to the plasma membrane. Our results thus identify two novel effectors of PtdIns(4,5)P(2) regulating cell growth and actin organization and suggest that Slm1 and Slm2 integrate inputs from the PtdIns(4,5)P(2) and TORC2 to modulate polarized actin assembly and growth.
Figures
References
-
- Abraham, R. T. (2002). Identification of TOR signaling complexes: more TORC for the cell growth engine. Cell 111, 9-12. - PubMed
-
- Alberts, A. S., Bouquin, N., Johnston, L. H., and Treisman, R. (1998). Analysis of RhoA-binding proteins reveals an interaction domain conserved in heterotrimeric G protein β subunits and the yeast response regulator protein Skn7. J. Biol. Chem. 273, 8616-8622. - PubMed
-
- Andrews, P. D., and Stark, M. J. (2000). Dynamic, Rho1p-dependent localization of Pkc1p to sites of polarized growth. J. Cell Sci. 113, 2685-2693. - PubMed
-
- Audhya, A., and Emr, S. D. (2002). Stt4 PI 4-kinase localizes to the plasma membrane and functions in the Pkc1-mediated MAP kinase cascade. Dev. Cell 2, 593-605. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
