

Protein folding: defining a "standard" set of experimental conditions and a preliminary kinetic data set of two-state proteins
- PMID: 15689503
- PMCID: PMC2279278
- DOI: 10.1110/ps.041205405
Protein folding: defining a "standard" set of experimental conditions and a preliminary kinetic data set of two-state proteins
Abstract
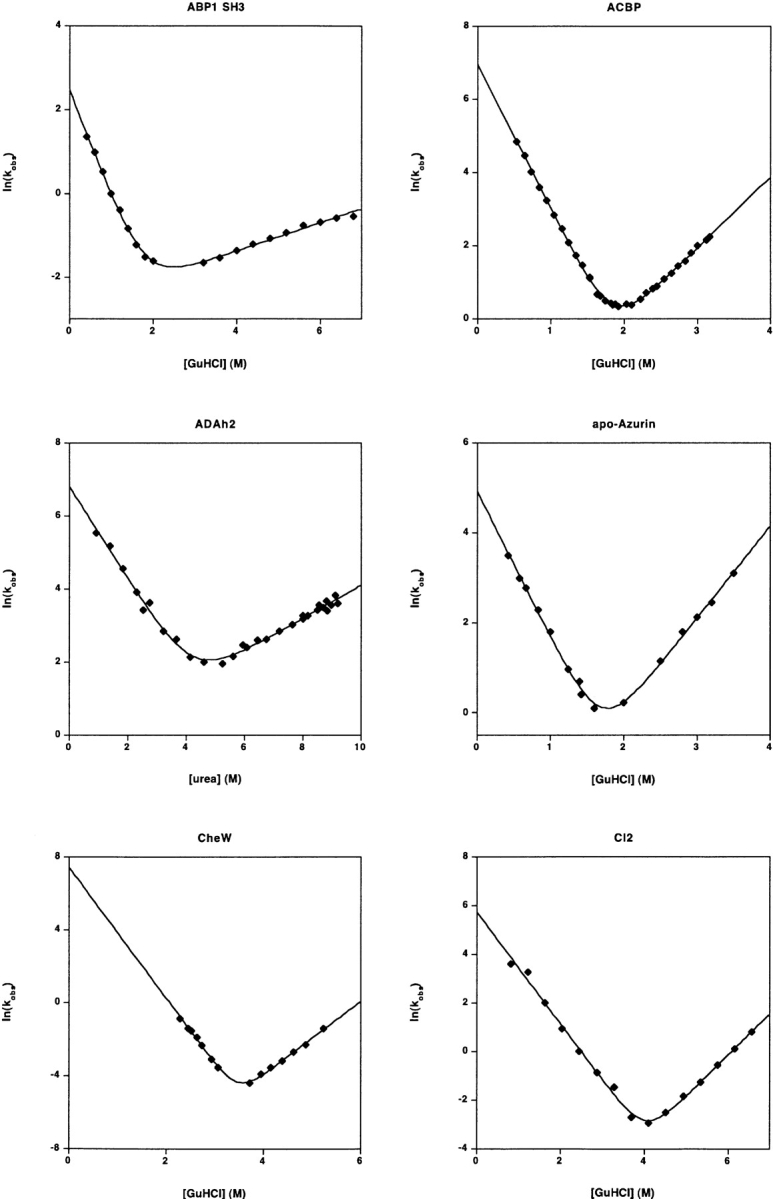
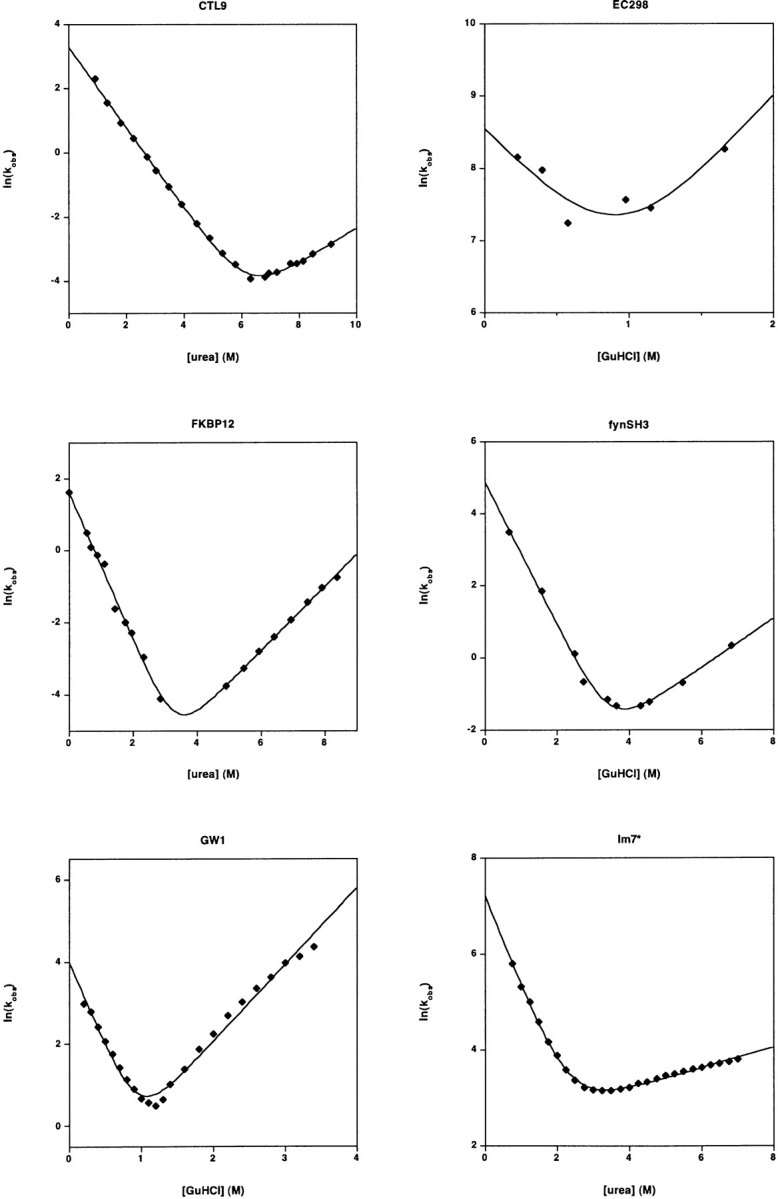
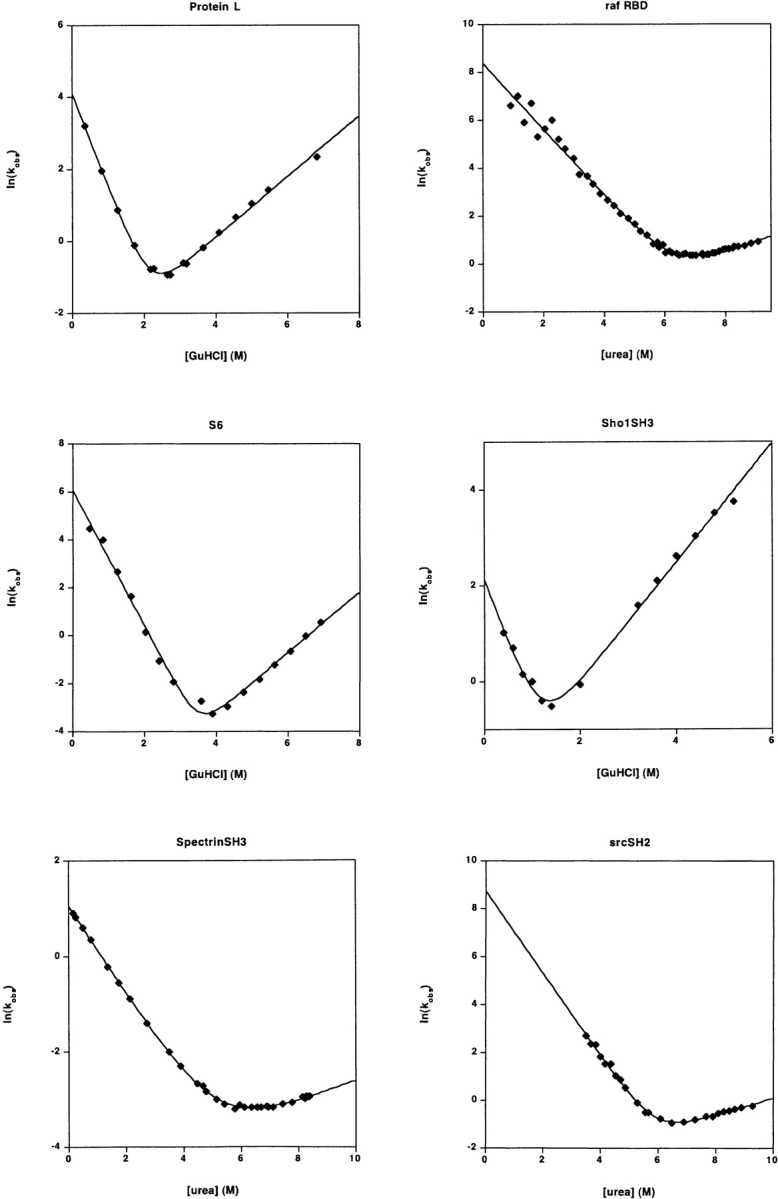
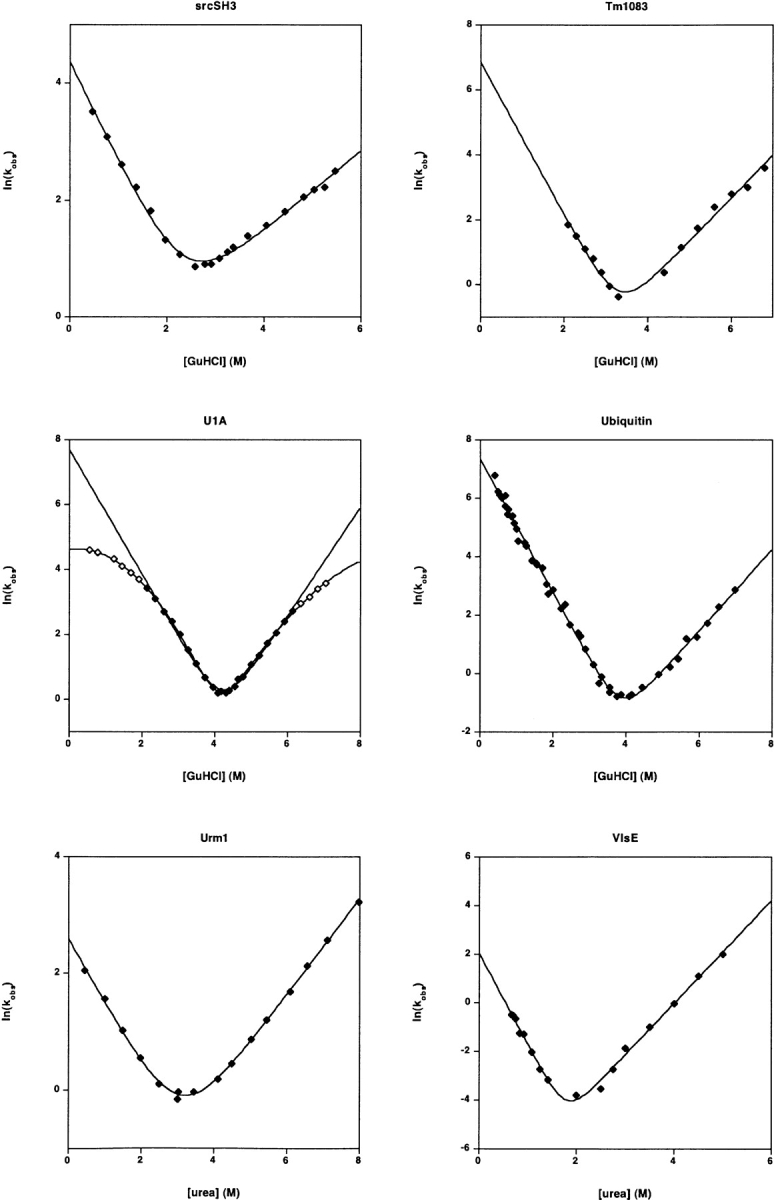
Recent years have seen the publication of both empirical and theoretical relationships predicting the rates with which proteins fold. Our ability to test and refine these relationships has been limited, however, by a variety of difficulties associated with the comparison of folding and unfolding rates, thermodynamics, and structure across diverse sets of proteins. These difficulties include the wide, potentially confounding range of experimental conditions and methods employed to date and the difficulty of obtaining correct and complete sequence and structural details for the characterized constructs. The lack of a single approach to data analysis and error estimation, or even of a common set of units and reporting standards, further hinders comparative studies of folding. In an effort to overcome these problems, we define here a "consensus" set of experimental conditions (25 degrees C at pH 7.0, 50 mM buffer), data analysis methods, and data reporting standards that we hope will provide a benchmark for experimental studies. We take the first step in this initiative by describing the folding kinetics of 30 apparently two-state proteins or protein domains under the consensus conditions. The goal of our efforts is to set uniform standards for the experimental community and to initiate an accumulating, self-consistent data set that will aid ongoing efforts to understand the folding process.
Figures
Similar articles
-
Folding of horse cytochrome c in the reduced state.J Mol Biol. 2001 Oct 5;312(5):1135-60. doi: 10.1006/jmbi.2001.4993. J Mol Biol. 2001. PMID: 11580255
-
High-pressure denaturation of staphylococcal nuclease proline-to-glycine substitution mutants.Biochemistry. 1996 Mar 26;35(12):3857-64. doi: 10.1021/bi952012g. Biochemistry. 1996. PMID: 8620010
-
Application of long-range order to predict unfolding rates of two-state proteins.Proteins. 2011 Mar;79(3):880-7. doi: 10.1002/prot.22925. Epub 2010 Dec 22. Proteins. 2011. PMID: 21287619
-
Topology, stability, sequence, and length: defining the determinants of two-state protein folding kinetics.Biochemistry. 2000 Sep 19;39(37):11177-83. doi: 10.1021/bi000200n. Biochemistry. 2000. PMID: 10985762 Review.
-
Cooperativity and protein folding rates.Curr Opin Struct Biol. 2010 Feb;20(1):11-5. doi: 10.1016/j.sbi.2009.12.013. Epub 2010 Jan 21. Curr Opin Struct Biol. 2010. PMID: 20093004 Review.
Cited by
-
Nonequilibrium single molecule protein folding in a coaxial mixer.Biophys J. 2008 Jul;95(1):352-65. doi: 10.1529/biophysj.107.127431. Epub 2008 Mar 13. Biophys J. 2008. PMID: 18339751 Free PMC article.
-
Methods for the accurate estimation of confidence intervals on protein folding phi-values.Protein Sci. 2006 Oct;15(10):2257-64. doi: 10.1110/ps.062230106. Protein Sci. 2006. PMID: 17008714 Free PMC article.
-
Thermodynamic and kinetic stability of a large multi-domain enzyme from the hyperthermophile Aeropyrum pernix.Extremophiles. 2010 Mar;14(2):213-23. doi: 10.1007/s00792-009-0300-0. Extremophiles. 2010. PMID: 20058042
-
Kinetic model of GPCR-G protein interactions reveals allokairic modulation of signaling.Nat Commun. 2022 Mar 8;13(1):1202. doi: 10.1038/s41467-022-28789-5. Nat Commun. 2022. PMID: 35260563 Free PMC article.
-
A Residue Quartet in the Extracellular Domain of the Prolactin Receptor Selectively Controls Mitogen-activated Protein Kinase Signaling.J Biol Chem. 2015 May 8;290(19):11890-904. doi: 10.1074/jbc.M115.639096. Epub 2015 Mar 17. J Biol Chem. 2015. PMID: 25784554 Free PMC article.
References
-
- Capaldi, A.P., Kleanthous, C., and Radford, S.E. 2002. Im7 folding mechanism: Misfolding on a path to the native state. Nat. Struct. Biol. 9 209–216. - PubMed
-
- Chiti, F., Taddei, N., van Nuland, N.A., Magherini, F., Stefani, M., Ramponi, G., and Dobson, C.M. 1998. Structural characterization of the transition state for folding of muscle acylphosphatase. J. Mol. Biol. 283 893–903. - PubMed
-
- de los Rios, M. A., and Plaxco, K.W. 2005. Apparent Debye-Huckel electrostatic effects in the folding of a simple, single domain protein. Biochemistry (in press). - PubMed
-
- Ferguson, N., Capaldi, A.P., James, R., Kleanthous, C., and Radford, S.E. 1999. Rapid folding with and without populated intermediates in the homologous four-helix proteins Im7 and Im9. J. Mol. Biol. 286 1597–1608. - PubMed
-
- Friel, C.T., Capaldi, A.P., and Radford, S.E. 2003. Structural analysis of the rate-limiting transition states in the folding of lm7 and lm9: Similarities and differences in the folding of homologous proteins. J. Mol. Biol. 326 293–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
