

The dynamic organization of gene-regulatory machinery in nuclear microenvironments
- PMID: 15689940
- PMCID: PMC1299247
- DOI: 10.1038/sj.embor.7400337
The dynamic organization of gene-regulatory machinery in nuclear microenvironments
Abstract
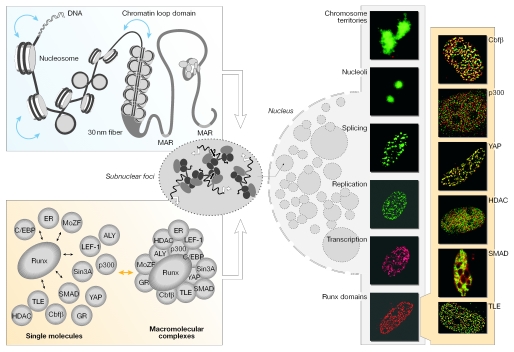
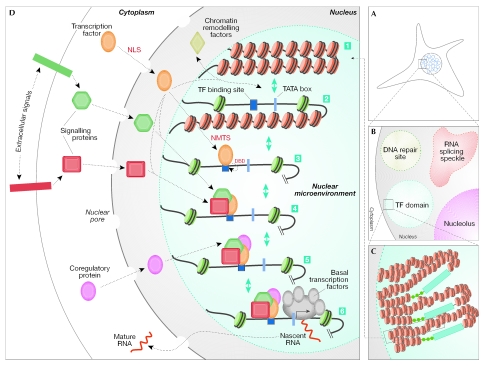
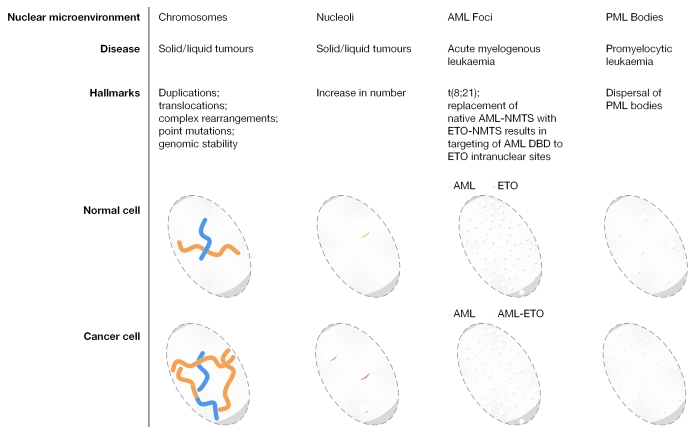
Nuclear components are functionally linked with the dynamic temporal and spatial compartmentalization, sorting and integration of regulatory information to facilitate its selective use. For example, the subnuclear targeting of transcription factors to punctate sites in the interphase nucleus mechanistically couples chromatin remodelling and the execution of signalling cascades that mediate gene expression with the combinatorial assembly of the regulatory machinery for biological control. In addition, a mitotic cycle of selective partitioning and sequential restoration of the transcriptional machinery provides a basis for the reassembly of regulatory complexes to render progeny cells competent for phenotypic gene expression. When this intranuclear targeting and localization of regulatory proteins is compromised, diseases, such as cancer, can occur. A detailed understanding of this process will provide further options for diagnosis and treatment.
Figures
References
-
- Baserga R, Peruzzi F, Reiss K (2003) The IGF-1 receptor in cancer biology. Int J Cancer 107: 873–877 - PubMed
-
- Berube NG, Jagla M, Smeenk C, De Repentigny Y, Kothary R, Picketts DJ (2002) Neurodevelopmental defects resulting from ATRX overexpression in transgenic mice. Hum Mol Genet 11: 253–261 - PubMed
-
- Brown KE, Baxter J, Graf D, Merkenschlager M, Fisher AG (1999) Dynamic repositioning of genes in the nucleus of lymphocytes preparing for cell division. Mol Cell 3: 207–217 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
