

5' CArG degeneracy in smooth muscle alpha-actin is required for injury-induced gene suppression in vivo
- PMID: 15690088
- PMCID: PMC546420
- DOI: 10.1172/JCI22648
5' CArG degeneracy in smooth muscle alpha-actin is required for injury-induced gene suppression in vivo
Abstract
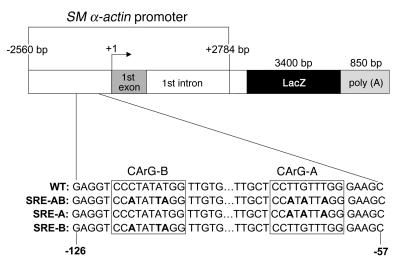
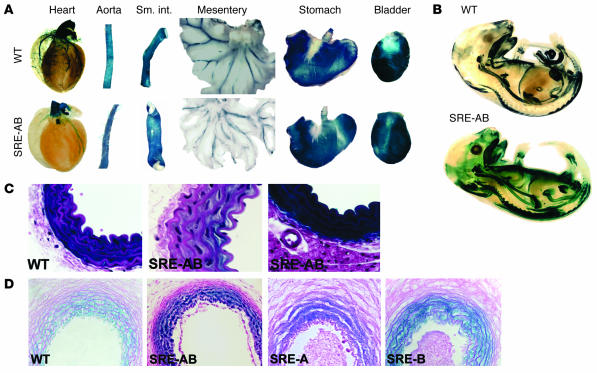
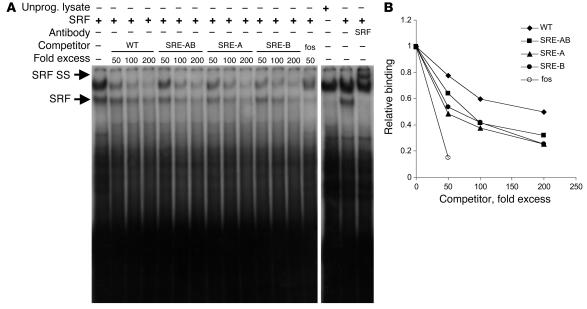
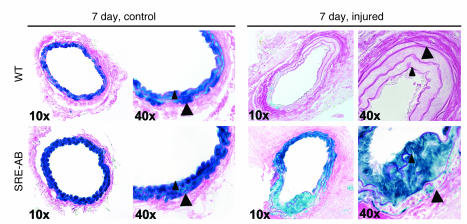
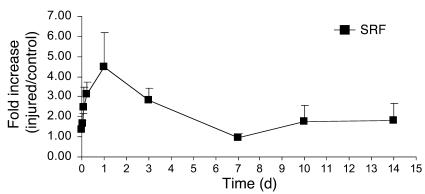
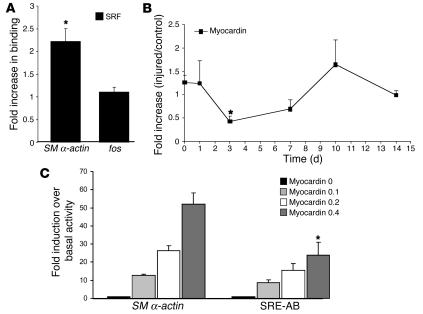
CC(A/T)6GG-dependent (CArG-dependent) and serum response factor-dependent (SRF-dependent) mechanisms are required for gene expression in smooth muscle cells (SMCs). However, an unusual feature of many SMC-selective promoter CArG elements is that they contain a conserved single G or C substitution in their central A/T-rich region, which reduces binding affinity for ubiquitously expressed SRF. We hypothesized that this CArG degeneracy contributes to cell-specific expression of smooth muscle alpha-actin in vivo, since substitution of c-fos consensus CArGs for the degenerate CArGs resulted in relaxed specificity in cultured cells. Surprisingly, our present results show that these substitutions have no effect on smooth muscle-specific transgene expression during normal development and maturation in transgenic mice. However, these substitutions significantly attenuated injury-induced downregulation of the mutant transgene under conditions where SRF expression was increased but expression of myocardin, a smooth muscle-selective SRF coactivator, was decreased. Finally, chromatin immunoprecipitation analyses, together with cell culture studies, suggested that myocardin selectively enhanced SRF binding to degenerate versus consensus CArG elements. Our results indicate that reductions in myocardin expression and the degeneracy of CArG elements within smooth muscle promoters play a key role in phenotypic switching of smooth muscle cells in vivo, as well as in mediating responses of CArG-dependent smooth muscle genes and growth regulatory genes under conditions in which these 2 classes of genes are differentially expressed.
Figures
Comment in
-
Defining smooth muscle cells and smooth muscle injury.J Clin Invest. 2005 Feb;115(2):221-4. doi: 10.1172/JCI24272. J Clin Invest. 2005. PMID: 15690076 Free PMC article. Review.
Similar articles
-
L-type voltage-gated Ca2+ channels modulate expression of smooth muscle differentiation marker genes via a rho kinase/myocardin/SRF-dependent mechanism.Circ Res. 2004 Aug 20;95(4):406-14. doi: 10.1161/01.RES.0000138582.36921.9e. Epub 2004 Jul 15. Circ Res. 2004. PMID: 15256479
-
Smooth muscle alpha-actin CArG elements coordinate formation of a smooth muscle cell-selective, serum response factor-containing activation complex.Circ Res. 2000 Feb 4;86(2):221-32. doi: 10.1161/01.res.86.2.221. Circ Res. 2000. PMID: 10666419
-
Substitution of the degenerate smooth muscle (SM) alpha-actin CC(A/T-rich)6GG elements with c-fos serum response elements results in increased basal expression but relaxed SM cell specificity and reduced angiotensin II inducibility.J Biol Chem. 1998 Apr 3;273(14):8398-406. doi: 10.1074/jbc.273.14.8398. J Biol Chem. 1998. PMID: 9525950
-
Serum response factor: toggling between disparate programs of gene expression.J Mol Cell Cardiol. 2003 Jun;35(6):577-93. doi: 10.1016/s0022-2828(03)00110-x. J Mol Cell Cardiol. 2003. PMID: 12788374 Review.
-
Control of smooth muscle development by the myocardin family of transcriptional coactivators.Curr Opin Genet Dev. 2004 Oct;14(5):558-66. doi: 10.1016/j.gde.2004.08.003. Curr Opin Genet Dev. 2004. PMID: 15380248 Review.
Cited by
-
Smooth muscle-specific genes are differentially sensitive to inhibition by Elk-1.Mol Cell Biol. 2005 Nov;25(22):9874-85. doi: 10.1128/MCB.25.22.9874-9885.2005. Mol Cell Biol. 2005. PMID: 16260603 Free PMC article.
-
MicroRNA26 attenuates vascular smooth muscle maturation via endothelial BMP signalling.PLoS Genet. 2019 May 15;15(5):e1008163. doi: 10.1371/journal.pgen.1008163. eCollection 2019 May. PLoS Genet. 2019. PMID: 31091229 Free PMC article.
-
Methamphetamine induces thoracic aortic aneurysm/dissection through C/EBPβ.Biochim Biophys Acta Mol Basis Dis. 2022 Sep 1;1868(9):166447. doi: 10.1016/j.bbadis.2022.166447. Epub 2022 May 25. Biochim Biophys Acta Mol Basis Dis. 2022. PMID: 35643386 Free PMC article.
-
Chromatin immunoprecipitation (ChIP): revisiting the efficacy of sample preparation, sonication, quantification of sheared DNA, and analysis via PCR.PLoS One. 2011;6(10):e26015. doi: 10.1371/journal.pone.0026015. Epub 2011 Oct 25. PLoS One. 2011. PMID: 22046253 Free PMC article.
-
Transforming growth factor-β1-induced transcript 1 protein, a novel marker for smooth muscle contractile phenotype, is regulated by serum response factor/myocardin protein.J Biol Chem. 2011 Dec 2;286(48):41589-41599. doi: 10.1074/jbc.M111.250878. Epub 2011 Oct 8. J Biol Chem. 2011. PMID: 21984848 Free PMC article.
References
-
- Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004;84:767–801. - PubMed
-
- Fatigati V, Murphy RA. Actin and tropomyosin variants in smooth muscles. Dependence on tissue type. J. Biol. Chem. 1984;259:14383–14388. - PubMed
-
- Frid MG, Shekhonin BV, Koteliansky VE, Glukhova MA. Phenotypic changes of human smooth muscle cells during development: late expression of heavy caldesmon and calponin. Dev. Biol. 1992;153:185–193. - PubMed
-
- Hungerford JE, Owens GK, Argraves WS, Little CD. Development of the aortic vessel wall as defined by vascular smooth muscle and extracellular matrix markers. Dev. Biol. 1996;178:375–392. - PubMed
-
- Mack CP, Owens GK. Regulation of smooth muscle alpha-actin expression in vivo is dependent on CArG elements within the 5′ and first intron promoter regions. Circ. Res. 1999;84:852–861. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
