

Review
doi: 10.1016/j.tibs.2004.12.008.
RNA-polymerase-I-directed rDNA transcription, life and works
Affiliations
- PMID: 15691654
- PMCID: PMC3858833
- DOI: 10.1016/j.tibs.2004.12.008
Item in Clipboard
Review
RNA-polymerase-I-directed rDNA transcription, life and works
Trends Biochem Sci.
2005 Feb.
Abstract
In the extensive network of interdependent biochemical processes required for cell growth and division, there is mounting evidence that ribosomal DNA transcription by RNA polymerase I (pol I) not only drives cell growth via its direct role in production of the ribosomal RNA (rRNA) component of the protein-synthesis machinery, but that it is also crucial in determining the fate of the cell. Considerable progress has been made in recent years towards understanding both the function of components of the pol I transcription machinery and how cells accomplish the tight control of pol I transcription, balancing the supply of rRNA with demand under different growth conditions.
Figures
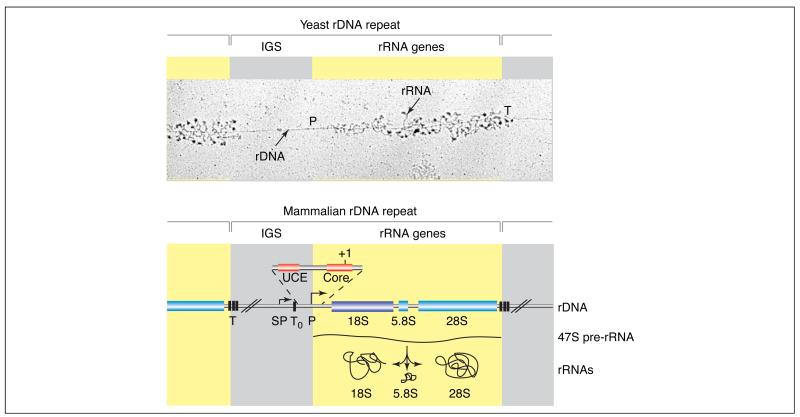
The repetitive nature of the rDNA is illustrated in an electron microscopic image of a yeast nuclear chromatin spread (‘Miller spread’). Progressively longer rRNAs (stained for associated proteins) emanate from the many pol I complexes as they transcribe the rDNA, beginning at the promoter (P) and finishing at the terminator (T). Beneath, a representative mammalian rDNA repeat is outlined (not drawn to scale). Each human rDNA repeat unit (GenBank accession number: U13369) of ~43 kb contains an intergenic spacer (IGS) of ~30 kb (grey), which contains the transcription regulatory elements, and ~13 kb of sequences encoding the precursor rRNA (yellow). The rRNA genes are present in a single transcription unit, transcribed by pol I to yield a 47S precursor rRNA that is, in part, co-transcriptionally processed and modified by methylation and pseudo-uridinylation to produce the mature 18S, 5.8S and 28S rRNAs. Pol I initiates transcription at the human rDNA promoter (P), which contains an essential core element from −45 to +18 relative to the start site (+1), and an upstream control element (UCE) from −156 to −107 [7]. A spacer-promoter (SP) upstream of the gene promoter directs pol-I-dependent transcription of short-lived transcripts of unknown function. Several transcription-termination elements are located at the 3′ end of the transcribed region of the rRNA genes (T) and immediately upstream of the rRNA gene transcription start site (T0), between the spacer and gene promoters [3].
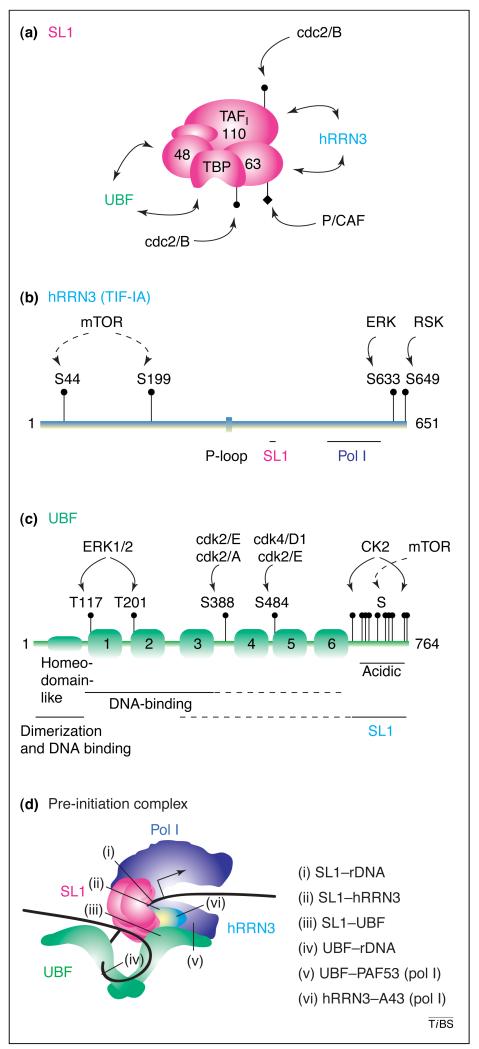
(a) Selectivity factor SL1 is an ~300-kDa complex of the TATA-binding protein (TBP) and at least three pol-I-specific TBP-associated factors (TAFIs) of 110, 63 and 48 kDa (GenBank accession numbers: NM_003194, NM_005679, NM_005680 and NM_005681). The mouse complex is TIF-IB and the homologous TAFIs are 95, 68 and 48 kDa, respectively (GenBank accession numbers: Y09974, Y09973 and Y09972). The SL1 subunits that interact with UBF and hRRN3 and post-translational modifications of TIF-IB [cdc2-cyclin B-mediated phosphorylation and p300/CBP-associated factor (P/CAF) acetylation] are indicated. (b) Human RRN3, a 651-amino acid polypeptide (GenBank accession number: NM_018427), and mouse homologue TIF-IA have sequence and functional homology to Saccharomyces cerevisiae Rrn3p. Conserved regions important for the interactions with SL1 (411–415) and Pol I (512–609) [18] and an ATP/GTP-binding site motif (P-loop; 333–340), of as yet unknown function, are indicated. TIF-IA activity can be regulated by the mTOR kinase and ERK-MAPK signalling pathways (ERK and RSK). (c) Human upstream binding factor UBF (also referred to as UBF1; GenBank accession number: NM_014233) is a 764-amino acid polypeptide of 97 kDa, which is highly conserved in vertebrates and is an activator of pol I transcription. The N-terminal domain is involved in dimerization (essential for the transcription activation function of UBF) and contains a homeo-domain-like motif (possibly involved in DNA binding). In addition, the protein has six high-mobility group (HMG) boxes (labelled 1–6). HMG boxes 1, 2 and 3 are involved in DNA binding through the minor groove. The C terminus, which contains 77% acidic amino acids and is rich in serine residues, can interact with SL1 and, in addition, might influence the binding of HMG box 1 to DNA in a UBF dimer. Human UBF2 is a 94-kDa protein of unknown function encoded by the same gene as UBF1, and is identical to UBF1 except for its lack of 37 amino acids from HMG box 2 due to alternative splicing [80]. Post-translational modifications of UBF include acetylation and phosphorylation via PI3K, cyclin-dependent kinases, the ERK–MAPK pathway and the mTOR pathway [signalling via p70 ribosomal S6 kinase to serine residues in the C terminus, also shown to be consensus phosphorylation sites for Casein Kinase 2 (CK2)]. (d) In the RNA polymerase I (pol I) pre-initiation complex (PIC), there are a multitude of protein–protein and protein–ribosomal promoter–DNA interactions (i) –(vi). For simplicity, only a single dimer of UBF contacting the rDNA is shown, but there might be a dimer at both the UCE and core sequences. UBF binds pol I, in part, through Pol-I-subunit PAF53, a homologue of yeast pol-I-subunit A49. Also involved in PIC formation, but of unknown identity, are TIF-IC, a mouse Pol-I-associated factor [41], and p70 [81]. The polymerase core subunits and pol-I-associated factors, other than hRRN3, are not detailed.
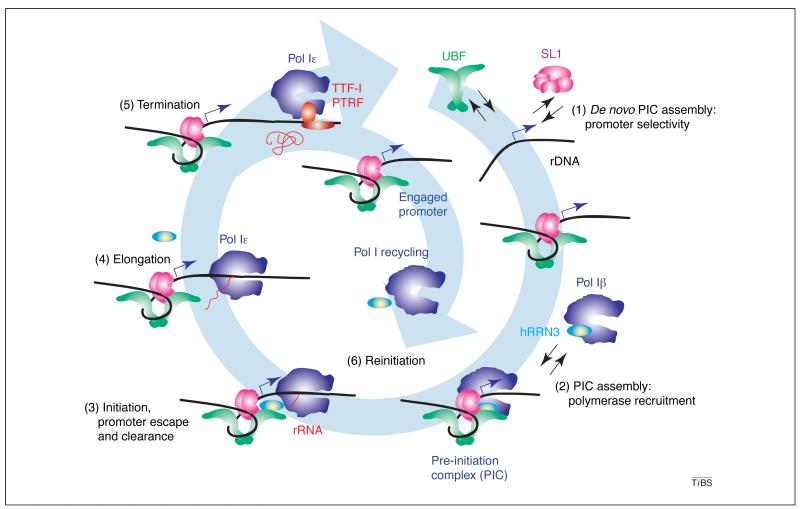
(1)
De novo PIC formation involves the selective binding of selectivity factor 1 (SL1) to the rDNA promoter, the incorporation of activator upstream binding protein (UBF) and (2) the recruitment of Pol Iβ by SL1. (3) Pol I initiates transcription upon promoter opening and, following promoter escape (3), pol I is converted into a processive enzyme (pol Iε), which elongates the nascent rRNA (4). (5) Transcription by pol I terminates at the 3′ end of the gene at specific sequences bound by termination factor TTF-I and transcript-release factor PTRF, with the concomitant release of pol I and the nascent rRNA. (6) SL1 and UBF remain promoter-bound following promoter clearance by pol I, and form a reinitiation scaffold onto which a pol Iβ complex, perhaps generated from recycled pol I and hRRN3, is recruited, and the resultant productive PIC can initiate another cycle of transcription.
References
-
- Grummt I. Life on a planet of its own: regulation of RNA polymerase I transcription in the nucleolus. Genes Dev. 2003;17:1691–1702. - PubMed
-
- Moss T, Stefanovsky VY. At the center of eukaryotic life. Cell. 2002;109:545–548. - PubMed
-
- Reeder RH. Regulation of RNA polymerase I transcription in yeast and vertebrates. Prog. Nucleic Acid Res. Mol. Biol. 1999;62:293–327. - PubMed
-
- Hannan KM, et al. Transcription by RNA polymerase I. Front. Biosci. 1998;3:d376–d398. - PubMed
-
- Comai L. Mechanism of RNA polymerase I transcription. Adv. Protein Chem. 2004;67:123–155. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
