

Multiple biological responses are induced by glycosylation-deficient hepatocyte growth factor
- PMID: 15693751
- PMCID: PMC1138963
- DOI: 10.1042/BJ20041698
Multiple biological responses are induced by glycosylation-deficient hepatocyte growth factor
Abstract
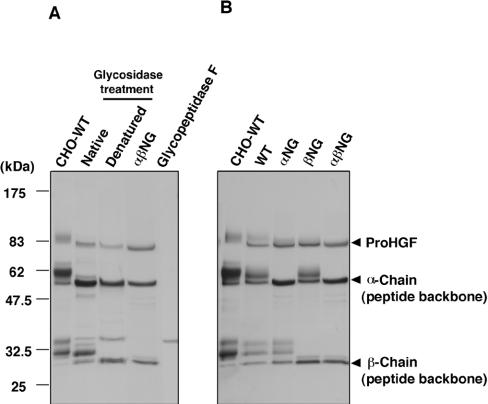
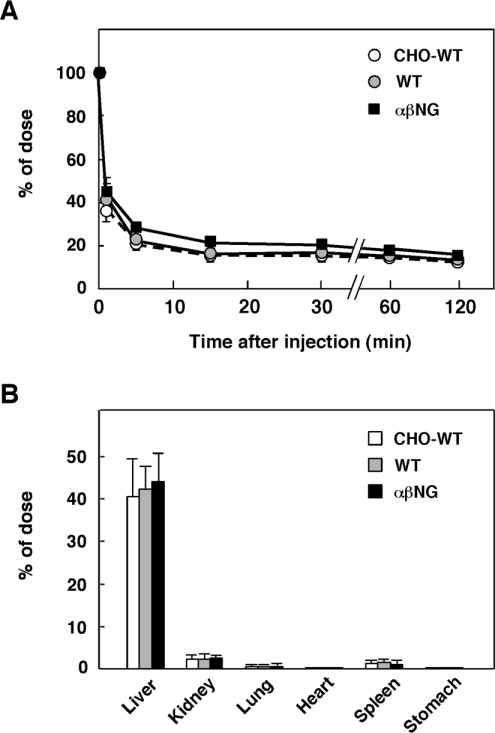
HGF (hepatocyte growth factor), a heterodimeric glycoprotein composed of alpha- and beta-chains, exerts biological activities through the c-Met receptor tyrosine kinase. The alpha-chain has three glycosylation sites, while the beta-chain has two; however, the role of sugar chains on HGF is still unknown. To address the significance of glycosylation of HGF, three different types of glycosylation-deficient HGFs, i.e. non-glycosylated in the alpha-chain, the beta-chain, and in both the alpha- and beta-chains, were respectively expressed in COS-7 cells and then purified from culture supernatants. Unexpectedly, glycosylation-deficient HGFs induced tyrosine phosphorylation of the c-Met receptor and subsequent phosphorylation of ERK (extracellular-signal-regulated kinase) and Akt in rat hepatocytes with the same potency as glycosylated HGF. Consistent with this, glycosylation-deficient HGFs strongly stimulated DNA synthesis of hepatocytes equal to glycosylated HGF. Likewise, glycosylation-deficient HGFs induced cell scattering and branching tubulogenesis in MDCK (Madin-Darby canine kidney) cells, and thus were indistinguishable from glycosylated HGF in biological activities. Glycosylation also did not affect stability, protease sensitivity and tissue distribution, although the plasma clearance of HGF was slightly prolonged by glycosylation deficiency. Glycosylation deficiency resulted in a decrease in post-transcriptional biosynthesis of HGF in the cells, whereas extracellularly secreted HGFs were efficiently activated to a two-chain form. These results indicate that glycosylation influences post-transcriptional biosynthesis of HGF, whereas biological activities and basic physicochemical characteristics are retained, even in completely non-glycosylated HGF. Hence, non-glycosylated HGF is promising as an alternative for glycosylated HGF in clinical applications.
Figures





Similar articles
-
SU5416 is a potent inhibitor of hepatocyte growth factor receptor (c-Met) and blocks HGF-induced invasiveness of human HepG2 hepatoma cells.J Hepatol. 2004 Aug;41(2):267-73. doi: 10.1016/j.jhep.2004.04.013. J Hepatol. 2004. PMID: 15288476
-
Structure, biosynthesis and biochemical properties of the HGF receptor in normal and malignant cells.EXS. 1993;65:131-65. EXS. 1993. PMID: 8380735 Review.
-
Hepatocyte growth factor enhances protein phosphatase Cdc25A inhibitor compound 5-induced hepatoma cell growth inhibition via Akt-mediated MAPK pathway.J Cell Physiol. 2005 Jun;203(3):510-9. doi: 10.1002/jcp.20243. J Cell Physiol. 2005. PMID: 15534860
-
Hepatocyte growth factor induces Wnt-independent nuclear translocation of beta-catenin after Met-beta-catenin dissociation in hepatocytes.Cancer Res. 2002 Apr 1;62(7):2064-71. Cancer Res. 2002. PMID: 11929826
-
Hepatocyte growth factor and its receptor, the tyrosine kinase encoded by the c-MET proto-oncogene.Cell Mol Biol (Noisy-le-grand). 1994 Jul;40(5):597-604. Cell Mol Biol (Noisy-le-grand). 1994. PMID: 7981617 Review.
Cited by
-
Distinct Localization of Mature HGF from its Precursor Form in Developing and Repairing the Stomach.Int J Mol Sci. 2019 Jun 17;20(12):2955. doi: 10.3390/ijms20122955. Int J Mol Sci. 2019. PMID: 31212972 Free PMC article.
-
Identification of mannose receptor as receptor for hepatocyte growth factor β-chain: novel ligand-receptor pathway for enhancing macrophage phagocytosis.J Biol Chem. 2012 Apr 13;287(16):13371-81. doi: 10.1074/jbc.M111.318568. Epub 2012 Feb 21. J Biol Chem. 2012. PMID: 22354962 Free PMC article.
-
Structural and Functional Insight Into the Glycosylation Impact Upon the HGF/c-Met Signaling Pathway.Front Cell Dev Biol. 2020 Jun 18;8:490. doi: 10.3389/fcell.2020.00490. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32626713 Free PMC article. Review.
-
Macrocyclic peptide-based inhibition and imaging of hepatocyte growth factor.Nat Chem Biol. 2019 Jun;15(6):598-606. doi: 10.1038/s41589-019-0285-7. Epub 2019 May 17. Nat Chem Biol. 2019. PMID: 31101918
-
Stable Ectopic Expression of ST6GALNAC5 Induces Autocrine MET Activation and Anchorage-Independence in MDCK Cells.PLoS One. 2016 Feb 5;11(2):e0148075. doi: 10.1371/journal.pone.0148075. eCollection 2016. PLoS One. 2016. PMID: 26848584 Free PMC article.
References
-
- Nakamura T., Nawa K., Ichihara A. Partial purification and characterization of hepatocyte growth factor from serum of hepatectomized rats. Biochem. Biophys. Res. Commun. 1984;122:1450–1459. - PubMed
-
- Nakamura T., Nishizawa T., Hagiya M., Seki T., Shimonishi M., Sugimura A., Tashiro K., Shimizu S. Molecular cloning and expression of human hepatocyte growth factor. Nature (London) 1989;342:440–443. - PubMed
-
- Matsumoto K., Nakamura T. Hepatocyte growth factor (HGF) as a tissue organizer for organogenesis and regeneration. Biochem. Biophys. Res. Commun. 1997;239:639–644. - PubMed
-
- Birchmeier C., Gherardi E. Developmental roles of HGF/SF and its receptor, the c-Met tyrosine kinase. Trends Cell Biol. 1998;8:404–410. - PubMed
-
- Bottaro D. P., Rubin J. S., Faletto D. L., Chan A. M., Kmiecik T. E., Vande Woude G. F., Aaronson S. A. Identification of the hepatocyte growth factor receptor as the c-met proto-oncogene product. Science. 1991;251:802–804. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous