

Live imaging of wound inflammation in Drosophila embryos reveals key roles for small GTPases during in vivo cell migration
- PMID: 15699212
- PMCID: PMC2171743
- DOI: 10.1083/jcb.200405120
Live imaging of wound inflammation in Drosophila embryos reveals key roles for small GTPases during in vivo cell migration
Abstract
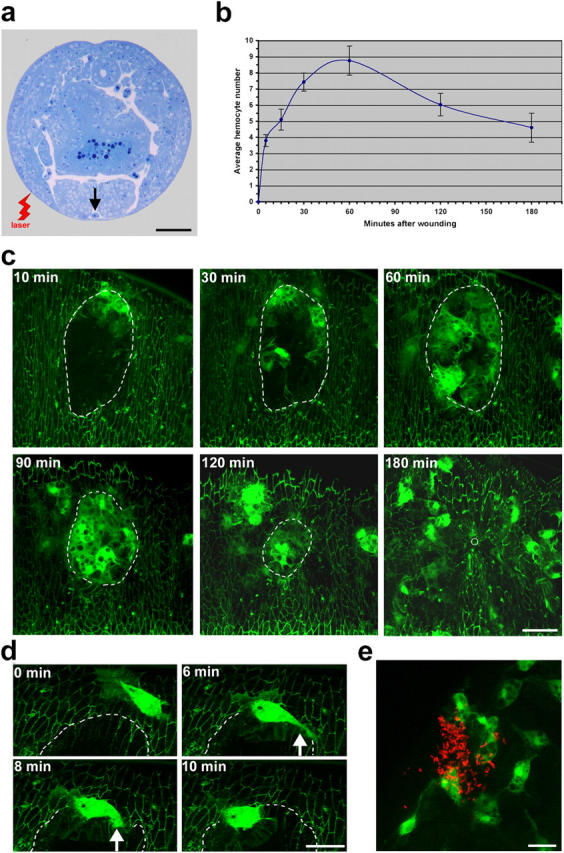
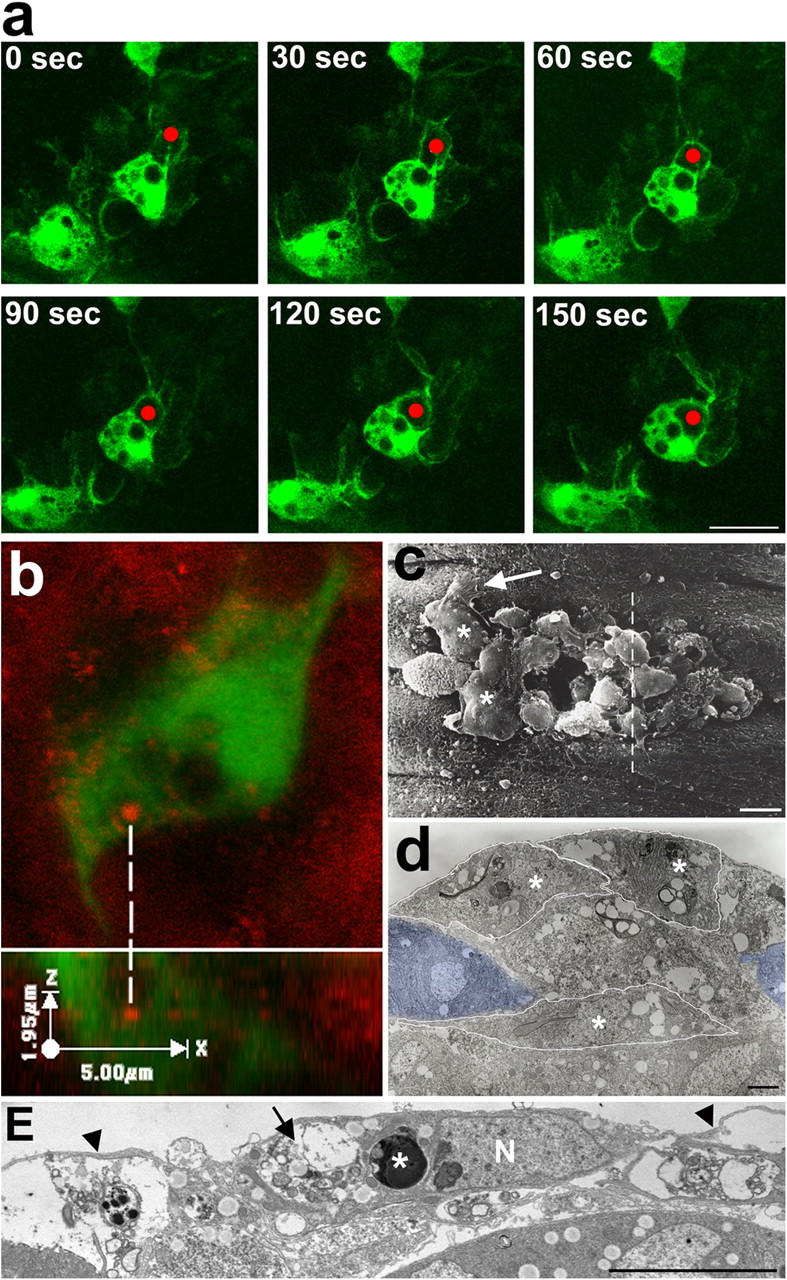
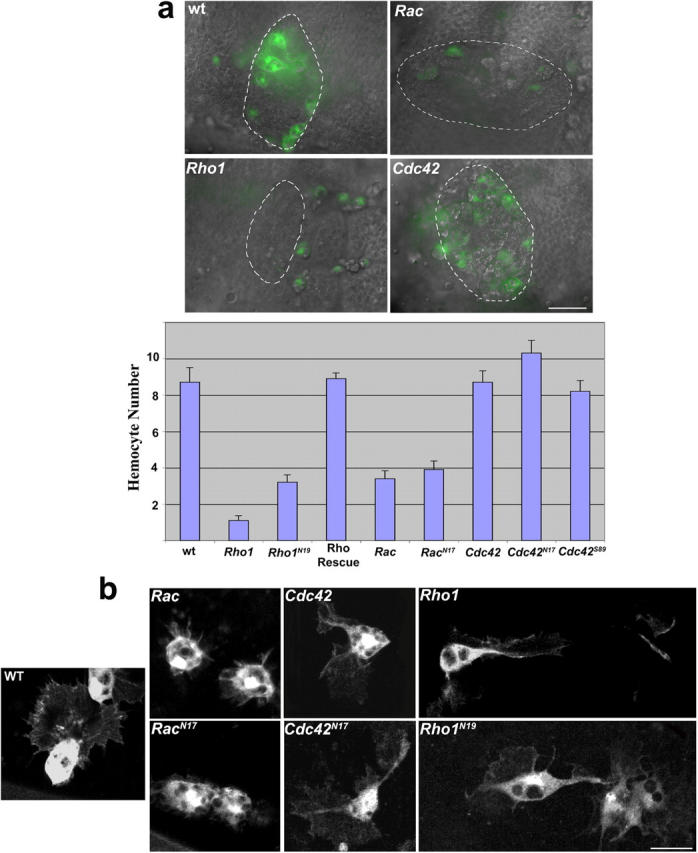
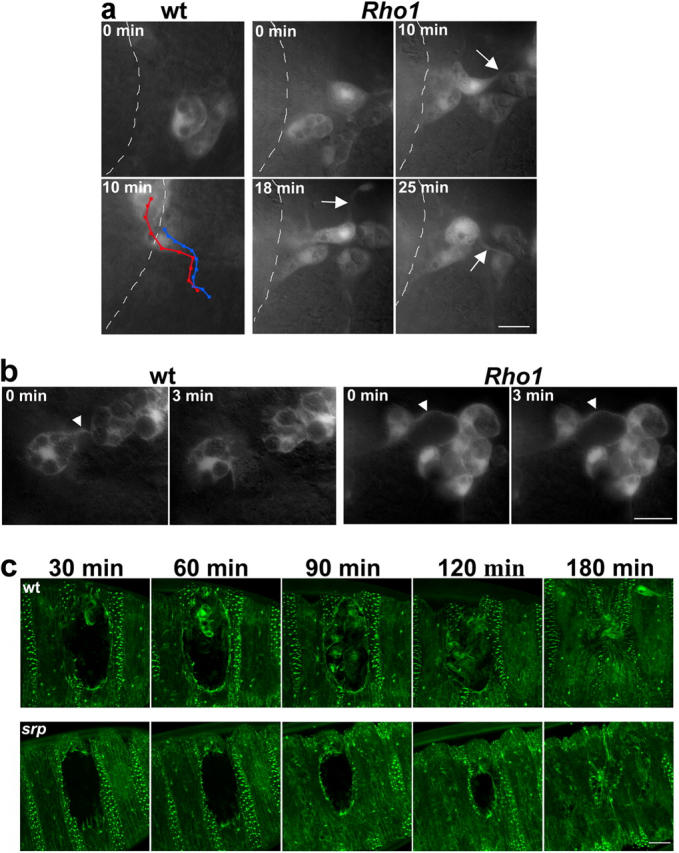
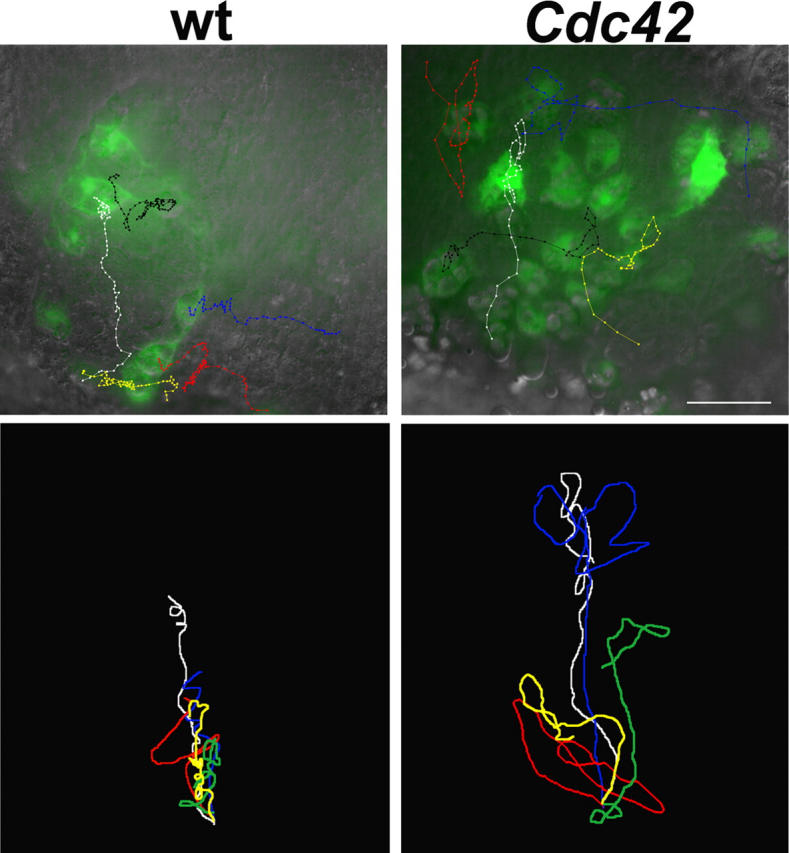
Aa robust inflammatory response to tissue damage and infection is conserved across almost all animal phyla. Neutrophils and macrophages, or their equivalents, are drawn to the wound site where they engulf cell and matrix debris and release signals that direct components of the repair process. This orchestrated cell migration is clinically important, and yet, to date, leukocyte chemotaxis has largely been studied in vitro. Here, we describe a genetically tractable in vivo wound model of inflammation in the Drosophila melanogaster embryo that is amenable to cinemicroscopy. For the first time, we are able to examine the roles of Rho-family small GTPases during inflammation in vivo and show that Rac-mediated lamellae are essential for hemocyte motility and Rho signaling is necessary for cells to retract from sites of matrix- and cell-cell contacts. Cdc42 is necessary for maintaining cellular polarity and yet, despite in vitro evidence, is dispensable for sensing and crawling toward wound cues.
Figures
References
-
- Cho, N.K., L. Keyes, E. Johnson, J. Heller, L. Ryner, F. Karim, and M.A. Krasnow. 2002. Developmental control of blood cell migration by the Drosophila VEGF pathway. Cell. 108:865–876. - PubMed
-
- Chou, J., N.A. Burke, A. Iwabu, S.C. Watkins, and A. Wells. 2003. Directional motility induced by epidermal growth factor requires Cdc42. Exp. Cell Res. 287:47–56. - PubMed
-
- Dovi, J.V., L.K. He, and L.A. DiPietro. 2003. Accelerated wound closure in neutrophil-depleted mice. J. Leukoc. Biol. 73:448–455. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
