

Plasmodium liver stage developmental arrest by depletion of a protein at the parasite-host interface
- PMID: 15699336
- PMCID: PMC548321
- DOI: 10.1073/pnas.0408442102
Plasmodium liver stage developmental arrest by depletion of a protein at the parasite-host interface
Abstract
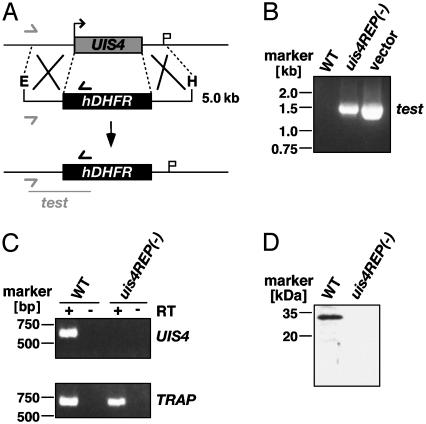
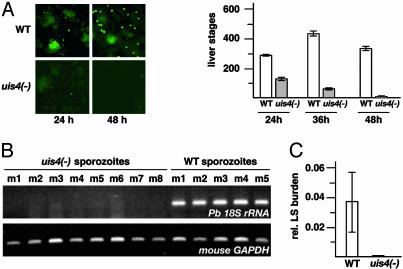
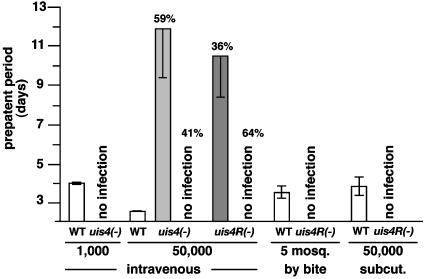
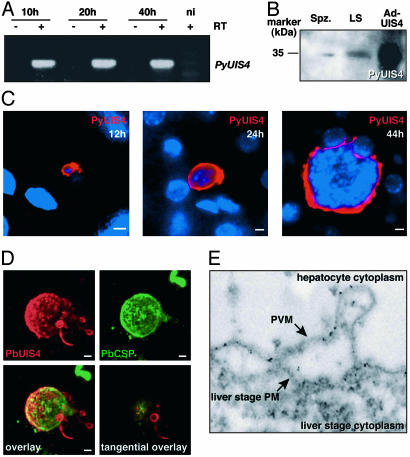
Plasmodium parasites of mammals, including the species that cause malaria in humans, infect the liver first and develop there into clinically silent liver stages. Liver stages grow and ultimately produce thousands of first-generation merozoites, which initiate the erythrocytic cycles causing malaria pathology. Here, we present a Plasmodium protein with a critical function for complete liver stage development. UIS4 (up-regulated in infective sporozoites gene 4) is expressed exclusively in infective sporozoites and developing liver stages, where it localizes to the parasitophorous vacuole membrane. Targeted gene disruption of UIS4 in the rodent model malaria parasite Plasmodium berghei generated knockout parasites that progress through the malaria life cycle until after hepatocyte invasion but are severely impaired in further liver stage development. Immunization with UIS4 knockout sporozoites completely protects mice against subsequent infectious WT sporozoite challenge. Genetically attenuated liver stages may thus induce immune responses, which inhibit subsequent infection of the liver with WT parasites.
Figures
References
-
- Kappe, S. H., Kaiser, K. & Matuschewski, K. (2003) Trends Parasitol. 19, 135-143. - PubMed
-
- Meis, J. F., Verhave, J. P., Jap, P. H., Sinden, R. E. & Meuwissen, J. H. (1983) Nature 302, 424-426. - PubMed
-
- Meis, J. F., Verhave, J. P., Jap, P. H. & Meuwissen, J. H. (1985) Cell Tissue Res. 241, 353-360. - PubMed
-
- Meis, J. F. & Verhave, J. P. (1988) Adv. Parasitol. 27, 1-61. - PubMed
-
- Shortt, H. E. & Garnham, P. C. C. (1948) Nature 161, 126. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
