

A high-throughput, near-saturating screen for type III effector genes from Pseudomonas syringae
- PMID: 15701698
- PMCID: PMC549004
- DOI: 10.1073/pnas.0409660102
A high-throughput, near-saturating screen for type III effector genes from Pseudomonas syringae
Abstract
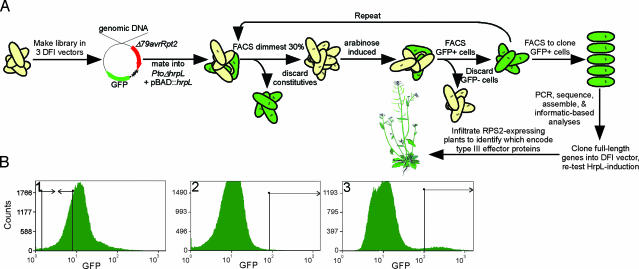
Pseudomonas syringae strains deliver variable numbers of type III effector proteins into plant cells during infection. These proteins are required for virulence, because strains incapable of delivering them are nonpathogenic. We implemented a whole-genome, high-throughput screen for identifying P. syringae type III effector genes. The screen relied on FACS and an arabinose-inducible hrpL sigma factor to automate the identification and cloning of HrpL-regulated genes. We determined whether candidate genes encode type III effector proteins by creating and testing full-length protein fusions to a reporter called Delta79AvrRpt2 that, when fused to known type III effector proteins, is translocated and elicits a hypersensitive response in leaves of Arabidopsis thaliana expressing the RPS2 plant disease resistance protein. Delta79AvrRpt2 is thus a marker for type III secretion system-dependent translocation, the most critical criterion for defining type III effector proteins. We describe our screen and the collection of type III effector proteins from two pathovars of P. syringae. This stringent functional criteria defined 29 type III proteins from P. syringae pv. tomato, and 19 from P. syringae pv. phaseolicola race 6. Our data provide full functional annotation of the hrpL-dependent type III effector suites from two sequenced P. syringae pathovars and show that type III effector protein suites are highly variable in this pathogen, presumably reflecting the evolutionary selection imposed by the various host plants.
Figures

Comment in
-
Powerful screens for bacterial virulence proteins.Proc Natl Acad Sci U S A. 2005 Mar 8;102(10):3527-8. doi: 10.1073/pnas.0500724102. Epub 2005 Feb 28. Proc Natl Acad Sci U S A. 2005. PMID: 15738387 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
