

Symmetry and frustration in protein energy landscapes: a near degeneracy resolves the Rop dimer-folding mystery
- PMID: 15701699
- PMCID: PMC548997
- DOI: 10.1073/pnas.0409572102
Symmetry and frustration in protein energy landscapes: a near degeneracy resolves the Rop dimer-folding mystery
Abstract
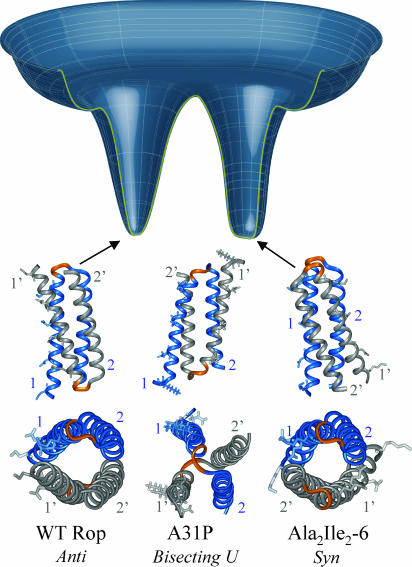
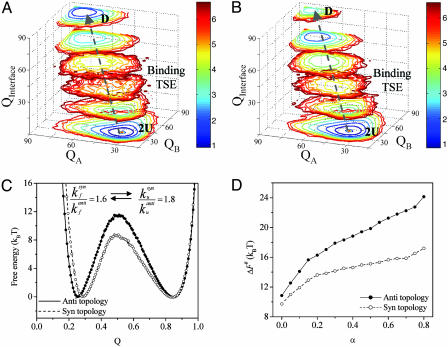
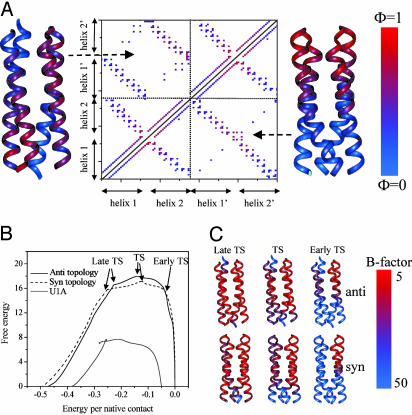
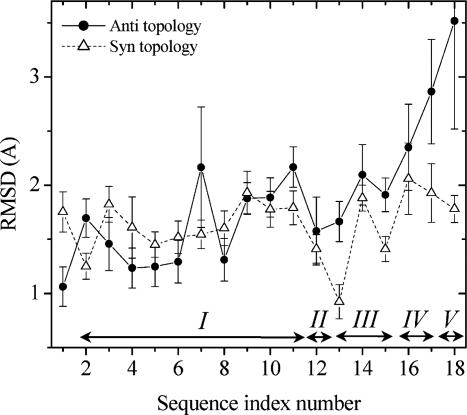
Protein folding has become one of the best understood biochemical reactions from a kinetic viewpoint. The funneled energy landscape, a consequence of the minimal frustration achieved by evolution in sequences, explains how most proteins fold efficiently and robustly to their functional structure and allows robust prediction of folding kinetics. The folding of Rop (repressor of primer) dimer is exceptional because some of its mutants with a redesigned hydrophobic core both fold and unfold much faster than the WT protein, which seems to conflict with a simple funneled energy landscape for which topology mainly determines the kinetics. We propose that the mystery of Rop folding can be unraveled by assuming a double-funneled energy landscape on which there are two basins that correspond to distinct but related topological structures. Because of the near symmetry of the molecule, mutations can cause a conformational switch to a nearly degenerate yet distinct topology or lead to a mixture of both topologies. The topology predicted to have the lower free-energy barrier height for folding was further found by all-atom modeling to give a better structural fit for those mutants with the extreme folding and unfolding rates. Thus, the non-Hammond effects can be understood within energy-landscape theory if there are in fact two different but nearly degenerate structures for Rop. Mutations in symmetric and regular structures may give rise to frustration and thus result in degeneracy.
Figures
References
-
- Onuchic, J. N., Luthey-Schulten, Z. & Wolynes, P. G. (1997) Annu. Rev. Phys. Chem. 48, 539-594. - PubMed
-
- Onuchic, J. N. & Wolynes, P. G. (2004) Curr. Opin. Struct. Biol. 14, 70-75. - PubMed
-
- Gunasekaran, K., Eyles, S. J., Hagler, A. T. & Gierasch, L. M. (2001) Curr. Opin. Struct. Biol. 2001, 83-93. - PubMed
-
- Murzin, A., Brenner, S., Hubbard, T. & Chothia, C. (1995) J. Mol. Biol. 247, 536-540. - PubMed
-
- Orengo, C., Jones, D. & Thornton, J. (1994) Nature 372, 631-634. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
