

Identification and characterization of circulating human transitional B cells
- PMID: 15701725
- PMCID: PMC1895038
- DOI: 10.1182/blood-2004-11-4284
Identification and characterization of circulating human transitional B cells
Abstract
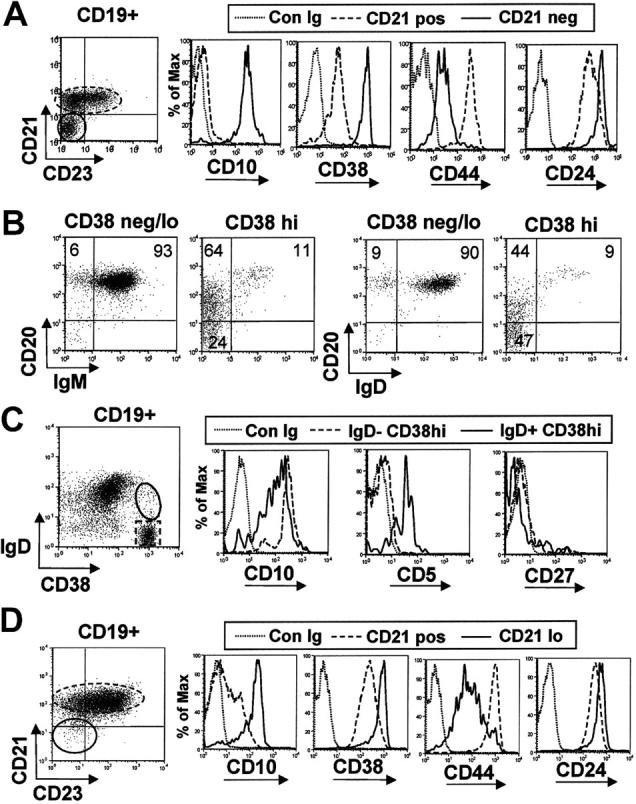
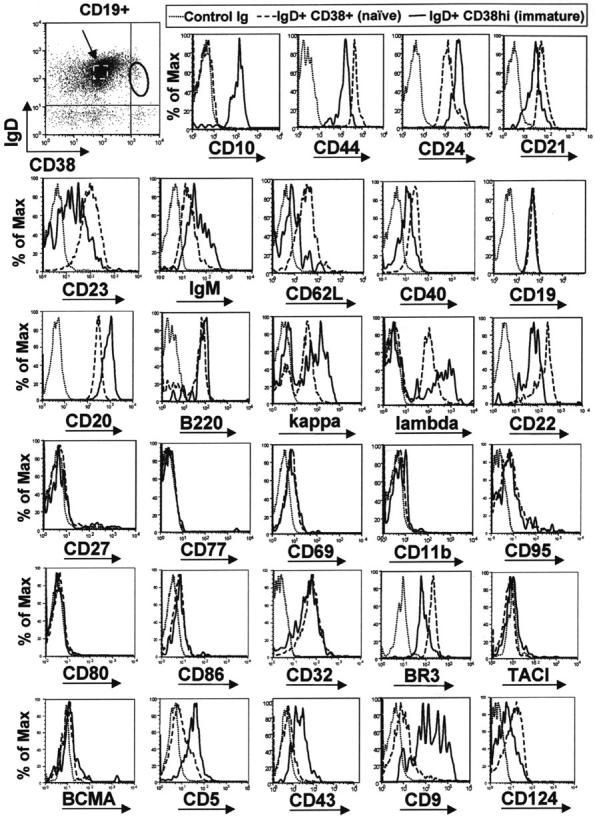
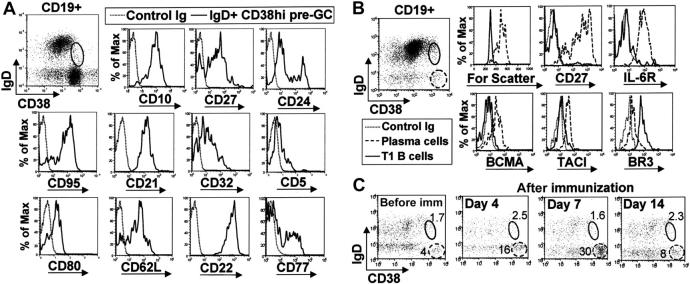
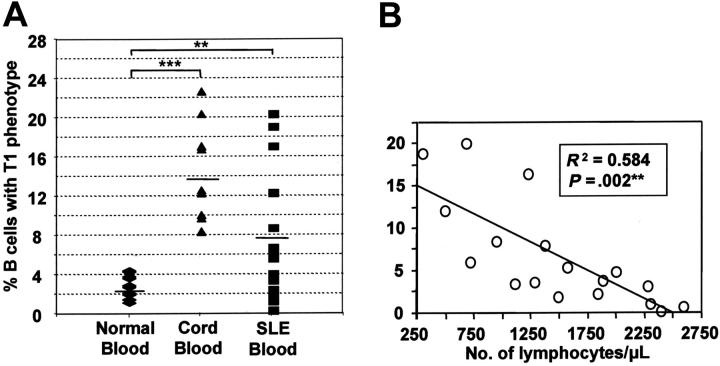
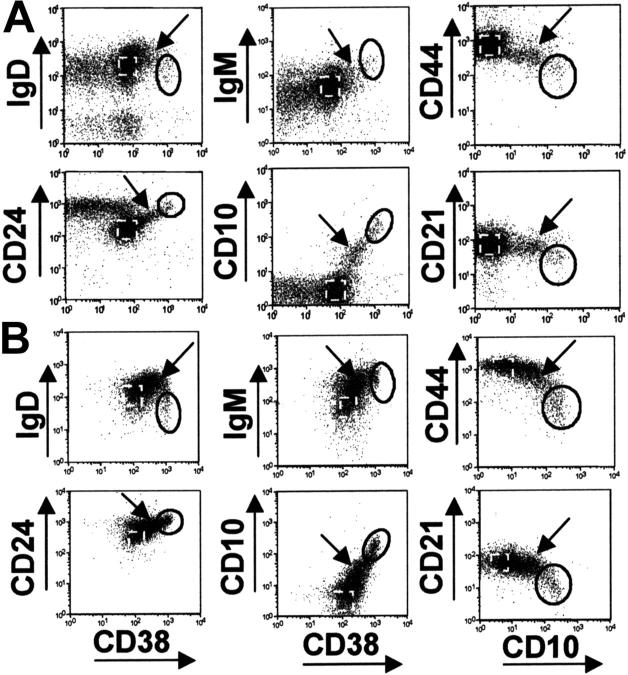
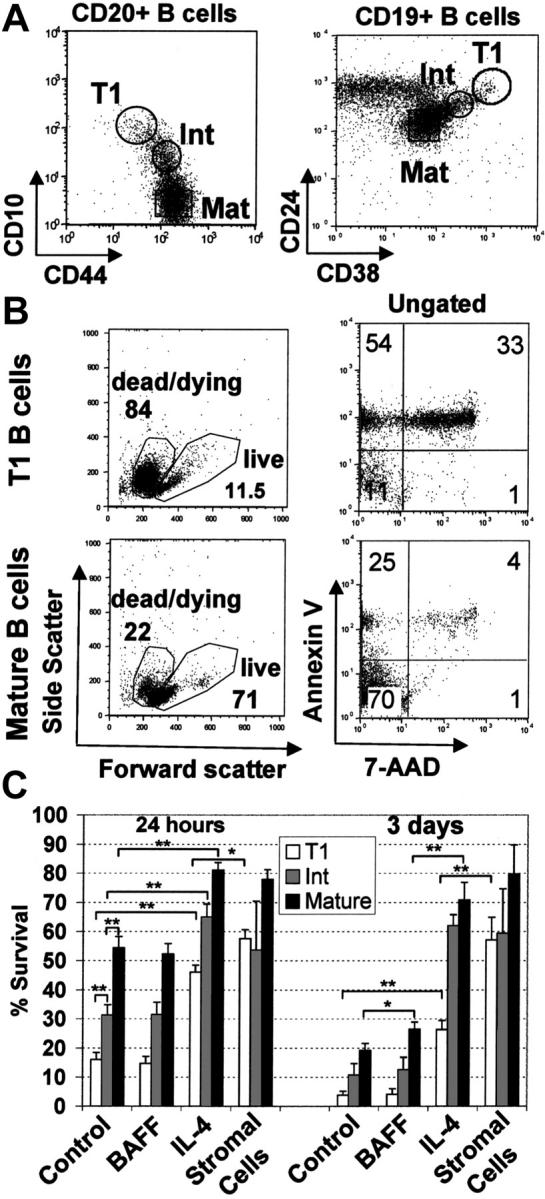
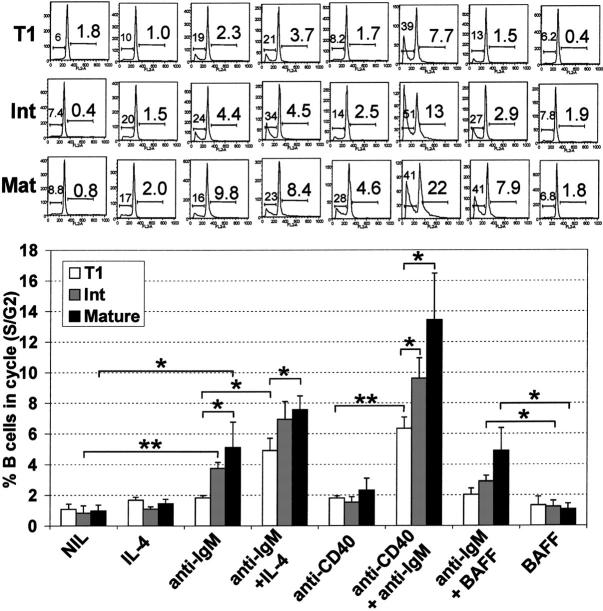
Murine B-cell development begins in bone marrow and results in the generation of immature transitional B cells that transit to the spleen to complete their maturation. It remains unclear whether the same developmental pathway takes place in humans. Using markers characteristic of human bone marrow immature B cells, we have identified a population of circulating human B cells with a phenotype most similar to mouse transitional type I (T1) B cells, although these human counterparts express CD5. These cells die rapidly in culture, and B-cell activation factor member of the tumor necrosis factor (TNF) family (BAFF) does not effect their survival regardless of B-cell receptor (BCR) stimulation. In contrast, bone marrow stromal cells or interleukin-4 (IL-4) significantly enhanced their survival. In the presence of T-cell signals provided by IL-4 or CD40 ligation, BCR stimulation can induce progression into cell cycle. Interestingly, circulating B cells that phenotypically and functionally resemble murine T2 B cells are found in cord blood and adult peripheral blood, suggesting that B-cell maturation may not be restricted to the spleen. Notably, increased proportions of T1 B cells were found in blood of patients with systemic lupus erythematosus (SLE), although bone marrow production and selection appeared to be normal.
Figures
References
-
- Hartley SB, Crosbie J, Brink R, Kantor AB, Basten A, Goodnow CC. Elimination from peripheral lymphoid-tissues of self-reactive lymphocytes-B recognizing membrane-bound antigens. Nature. 1991;353: 765-769. - PubMed
-
- Nemazee DA, Burki K. Clonal deletion of lymphocyte-B in a transgenic mouse bearing anti-Mhc class-I antibody genes. Nature. 1989;337: 562-566. - PubMed
-
- Norvell A, Mandik L, Monroe JG. Engagement of the antigen-receptor on immature murine B-lymphocytes results in death by apoptosis. J Immunol. 1995;154: 4404-4413. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
