

The p80 homology region of TEP1 is sufficient for its association with the telomerase and vault RNAs, and the vault particle
- PMID: 15701761
- PMCID: PMC549401
- DOI: 10.1093/nar/gki234
The p80 homology region of TEP1 is sufficient for its association with the telomerase and vault RNAs, and the vault particle
Abstract
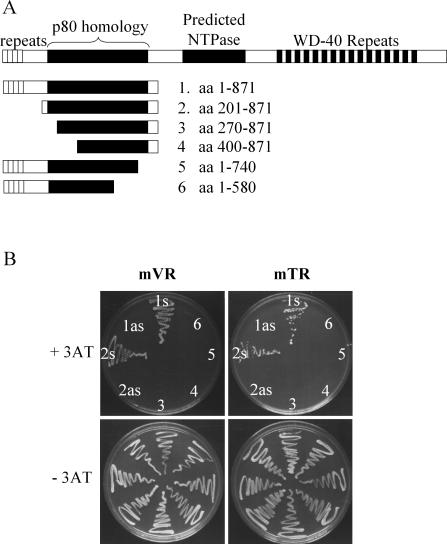
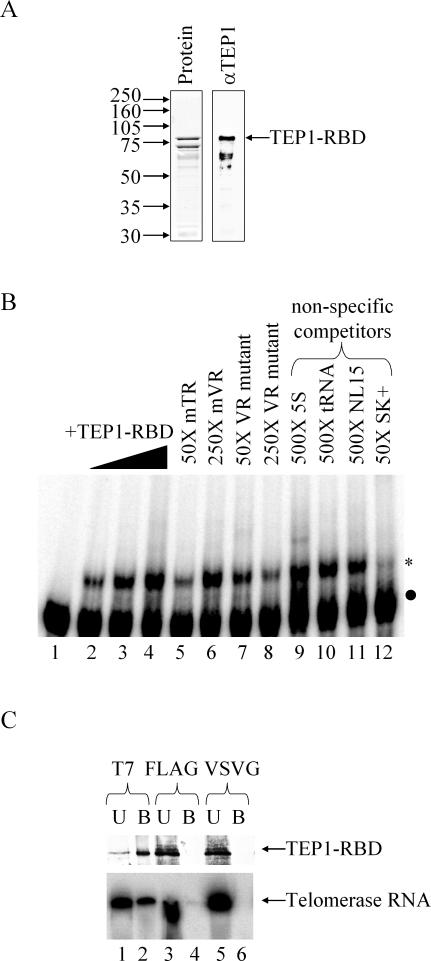
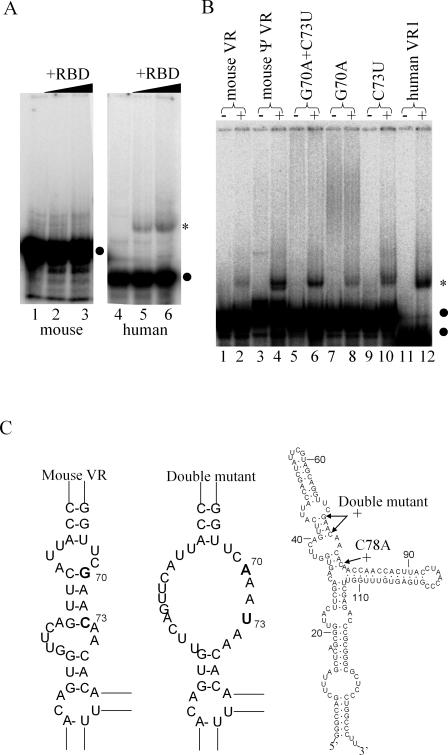
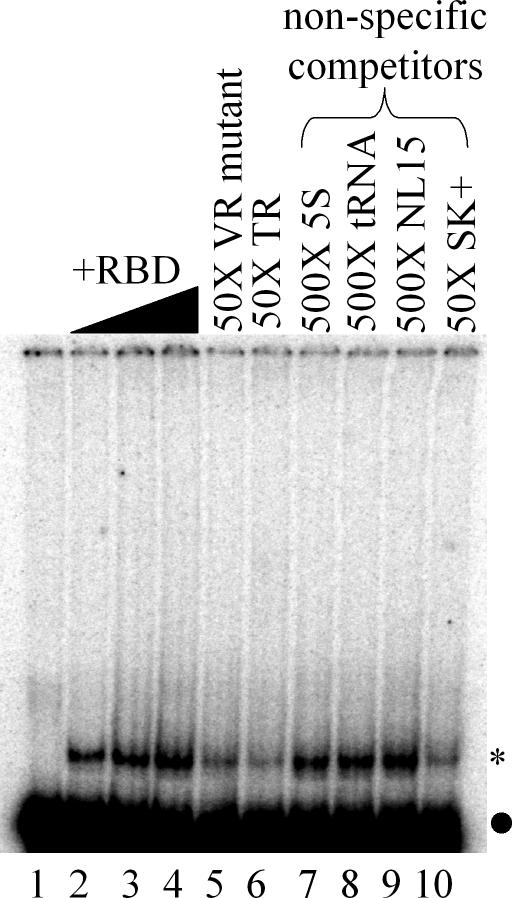
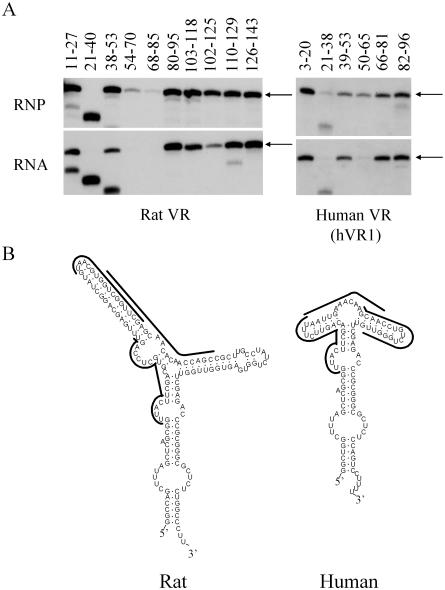
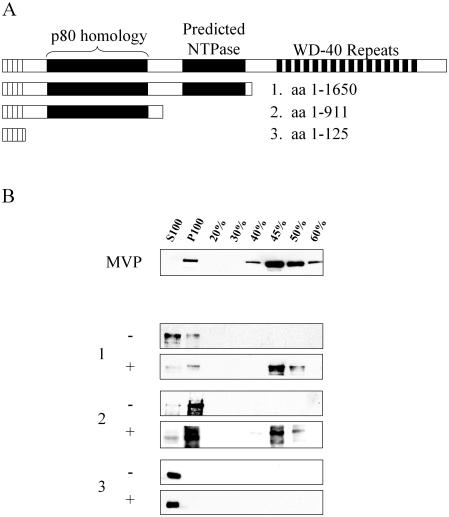
TEP1 is a protein component of two ribonucleoprotein complexes: vaults and telomerase. The vault-associated small RNA, termed vault RNA (VR), is dependent upon TEP1 for its stable association with vaults, while the association of telomerase RNA with the telomerase complex is independent of TEP1. Both of these small RNAs have been shown to interact with amino acids 1-871 of TEP1 in an indirect yeast three-hybrid assay. To understand the determinants of TEP1-RNA binding, we generated a series of TEP1 deletions and show by yeast three-hybrid assay that the entire Tetrahymena p80 homology region of TEP1 is required for its interaction with both telomerase and VRs. This region is also sufficient to target the protein to the vault particle. Electrophoretic mobility shift assays using the recombinant TEP1 RNA-binding domain (TEP1-RBD) demonstrate that it binds RNA directly, and that telomerase and VRs compete for binding. VR binds weakly to TEP1-RBD in vitro, but mutation of VR sequences predicted to disrupt helices near its central loop enhances binding. Antisense oligonucleotide-directed RNase H digestion of endogenous VR indicates that this region is largely single stranded, suggesting that TEP1 may require access to the VR central loop for efficient binding.
Figures
References
-
- Kong L.B., Siva A.C., Rome L.H., Stewart P.L. Structure of the vault, a ubiquitous cellular component. Structure Fold. Des. 1999;7:371–379. - PubMed
-
- Stephen A.G., Raval-Fernandes S., Huynh T., Torres M., Kickhoefer V.A., Rome L.H. Assembly of vault-like particles in insect cells expressing only the major vault protein. J. Biol. Chem. 2001;276:23217–23220. - PubMed
-
- Chugani D.C., Rome L.H., Kedersha N.L. Evidence that vault ribonucleoprotein particles localize to the nuclear pore complex. J. Cell. Sci. 1993;106:23–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
