

Neurosteroid administration and withdrawal alter GABAA receptor kinetics in CA1 hippocampus of female rats
- PMID: 15705652
- PMCID: PMC1464432
- DOI: 10.1113/jphysiol.2004.077297
Neurosteroid administration and withdrawal alter GABAA receptor kinetics in CA1 hippocampus of female rats
Abstract
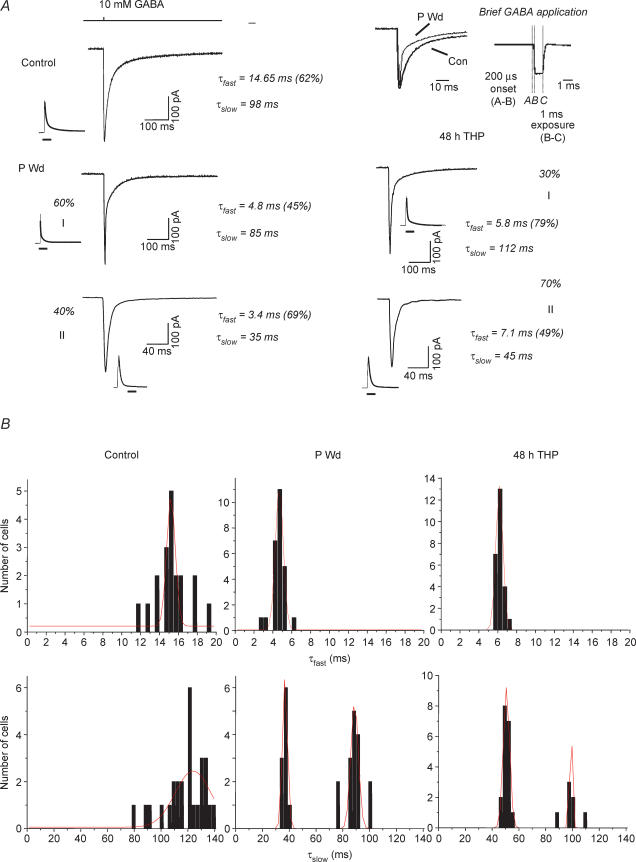
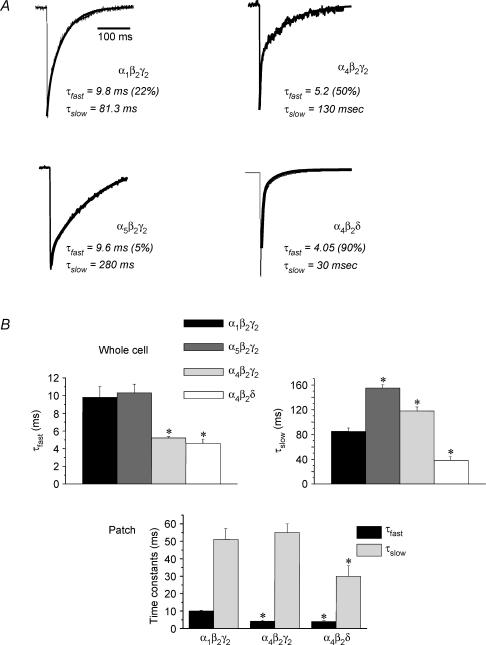
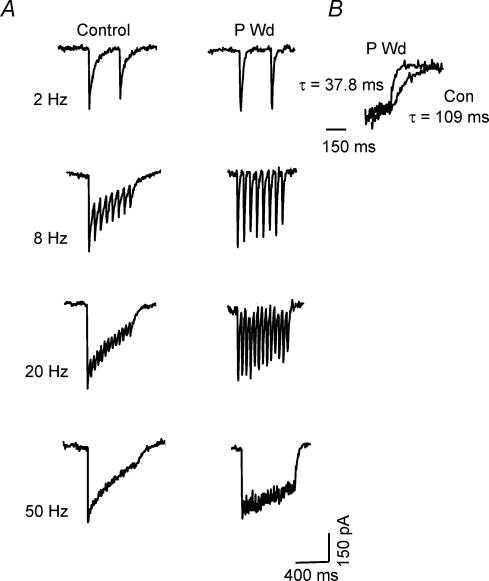
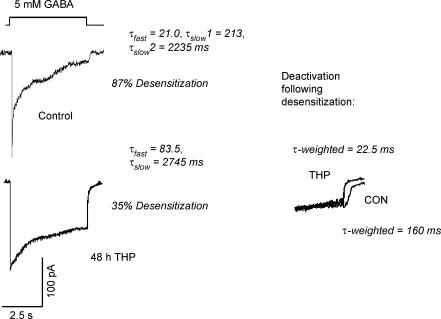
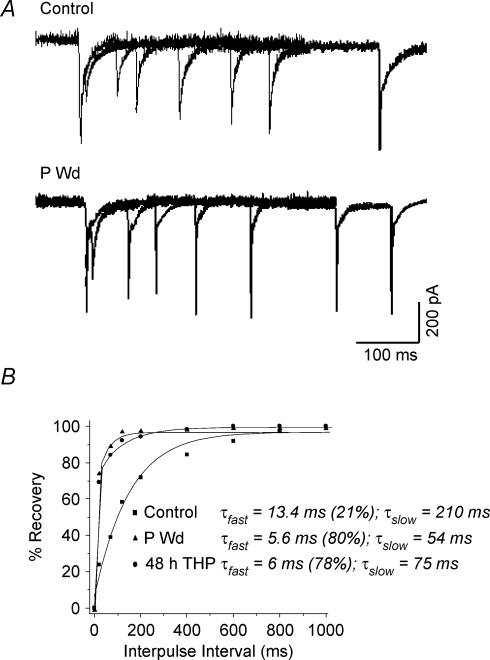
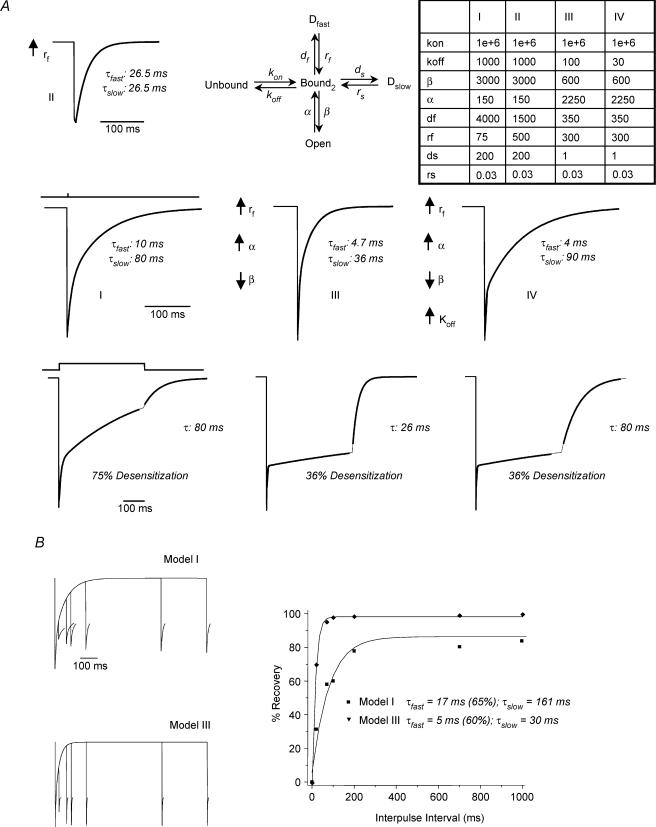
Withdrawal from the GABA-modulatory steroid 3alpha-OH-5alpha-pregnan-20-one (3alpha,5alpha-THP) following exposure of female rats to the parent compound progesterone (P) produces a syndrome characterized by behavioural excitability in association with up-regulation of the alpha4 subunit of the GABA(A) receptor (GABAR) in the hippocampus. Similar changes are seen after 48 h exposure to its stereoisomer, 3alpha,5beta-THP. Here, we further characterize the effects of P withdrawal on GABAR kinetics, using brief (1 ms) application of 5-10 mm GABA to outside-out patches from acutely isolated CA1 hippocampal pyramidal cells. Under control conditions, GABA-gated current deactivated biexponentially, with tau(fast) = 12-19 ms (45-60% of the current), and tau(slow) = 80-140 ms. P withdrawal resulted in marked acceleration of deactivation (tau(fast) = 3-7 ms and tau(slow) = 30-100 ms), as did 48 h exposure to 3alpha,5beta-THP (tau(fast) = 5-8 ms; tau(slow) = 40-120 ms). When recombinant receptors were tested in HEK-293 cells, a similar acceleration in tau(fast) was observed for alpha4beta2delta and alpha4beta2gamma2 GABARs, compared to alpha1beta2gamma2 and alpha5beta2gamma2 receptors. In addition, tau(slow) was also accelerated for alpha4beta2delta receptors, which are increased following steroid withdrawal. As predicted by the Jones-Westbrook model, this change was accompanied by reduced receptor desensitization as well as an acceleration of the rate of recovery from rapid desensitization. A theoretical analysis of the data suggested that steroid treatment leads to receptors with a greater stability of the bound, activatable state. This was achieved by altering multiple parameters, including desensitization and gating rates, within the model. These results suggest that fluctuations in endogenous steroids result in altered GABAR kinetics which may regulate neuronal excitability.
Figures
References
-
- Backstrom T. Epileptic seizures in women related to plasma estrogen and progesterone during the menstrual cycle. Acta Neurol Scand. 1976;54:321–347. - PubMed
-
- Bai D, Zhu G, Pennefather P, Jackson MF, Macdonald JF, Orser BA. Distinct functional and pharmacological properties of tonic and quantal inhibitory postsynaptic currents mediated by γ-aminobutyric acidA receptors in hippocampal neurons. Mol Pharmacol. 2001;59:814–824. - PubMed
-
- Banks MI, Pearce RA. Dual actions of volatile anesthetics on GABAA IPSCs: dissociation of blocking and prolonging effects. J Neurosci. 1999;90:120–134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous