

Tula hantavirus triggers pro-apoptotic signals of ER stress in Vero E6 cells
- PMID: 15708603
- PMCID: PMC7173054
- DOI: 10.1016/j.virol.2005.01.002
Tula hantavirus triggers pro-apoptotic signals of ER stress in Vero E6 cells
Abstract
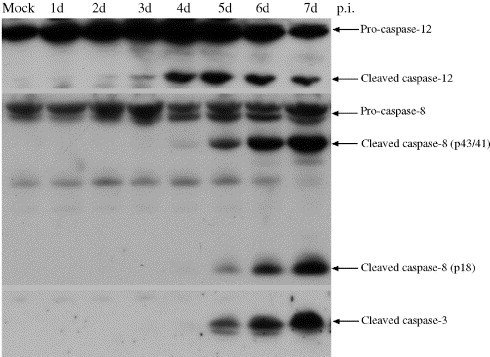
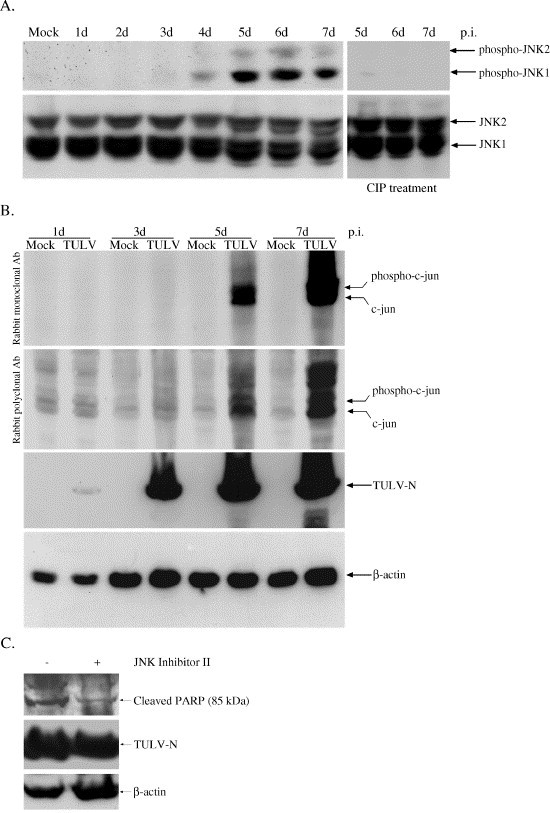
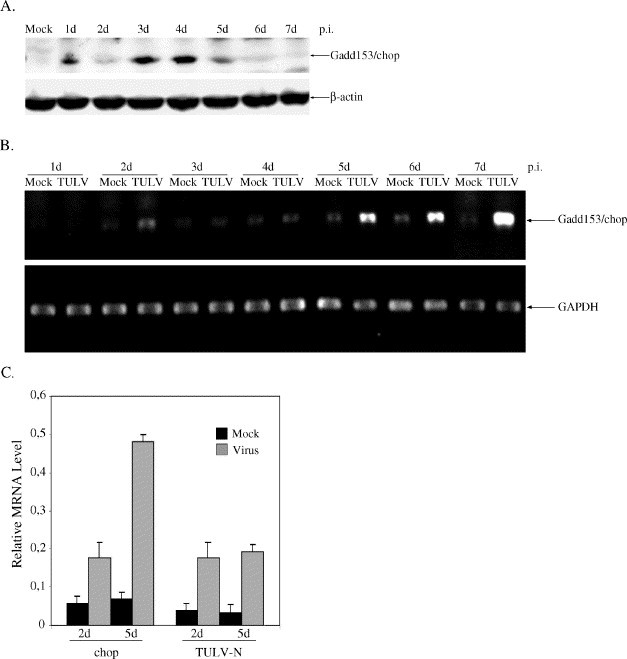
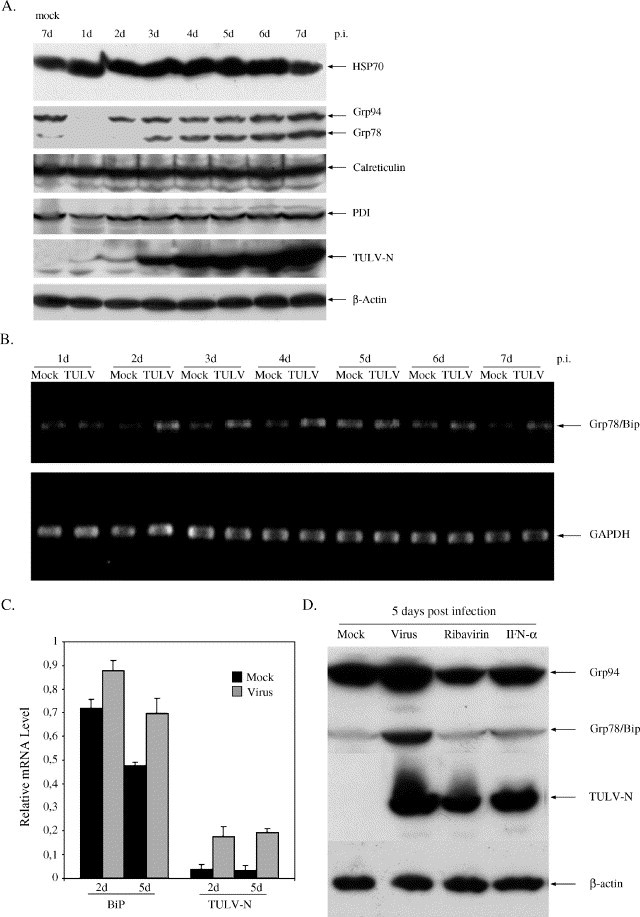
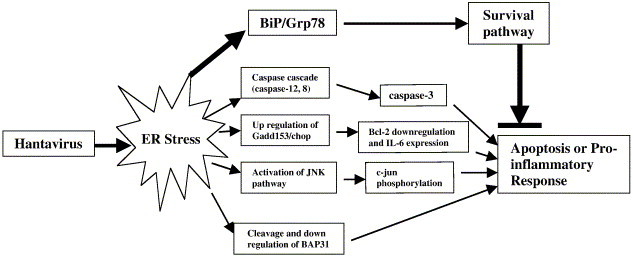
Tula virus is a member of the Hantavirus genus of the family Bunyaviridae. Viruses of this family have an unusual pattern of intracellular maturation at the ER-Golgi compartment. We recently found that Tula virus, similar to several other hantaviruses, is able to induce apoptosis in cultured cells [Li, X.D., Kukkonen, S., Vapalahti, O., Plyusnin, A., Lankinen, H., Vaheri, A., 2004. Tula hantavirus infection of Vero E6 cells induces apoptosis involving caspase 8 activation. J. Gen. Virol. 85, 3261-3268.]. However, the cellular mechanisms remain to be clarified. In this study, we demonstrate that the progressive replication of Tula virus in Vero E6 cells initiates several death programs that are intimately associated with ER stress: (1) early activation of ER-resident caspase-12; (2) phosphorylation of Jun NH2-terminal kinase (JNK) and its downstream target transcriptional factor, c-jun; (3) induction of the pro-apoptotic transcriptional factor, growth arrest- and DNA damage-inducible gene 153, or C/EBP homologous protein (Gadd153/chop); and (4) changes in the ER-membrane protein BAP31 implying cross-talk with the mitochondrial apoptosis pathway. Furthermore, we confirmed that a sustained ER stress was induced marked by an increased expression of an ER chaperone Grp78/BiP. Taken together, we have identified involvement of ER stress-mediated death program in Tula virus-infected Vero E6 cells which provides a new approach to understand the mechanisms in hantavirus-induced apoptosis.
Figures
Similar articles
-
Hantaviruses and TNF-alpha act synergistically to induce ERK1/2 inactivation in Vero E6 cells.Virol J. 2008 Sep 29;5:110. doi: 10.1186/1743-422X-5-110. Virol J. 2008. PMID: 18822184 Free PMC article.
-
Tula hantavirus infection of Vero E6 cells induces apoptosis involving caspase 8 activation.J Gen Virol. 2004 Nov;85(Pt 11):3261-3268. doi: 10.1099/vir.0.80243-0. J Gen Virol. 2004. PMID: 15483239
-
HFRS causing hantaviruses do not induce apoptosis in confluent Vero E6 and A-549 cells.J Med Virol. 2005 Jun;76(2):234-40. doi: 10.1002/jmv.20347. J Med Virol. 2005. PMID: 15834879
-
Proteasome inhibitor MG132-induced apoptosis via ER stress-mediated apoptotic pathway and its potentiation by protein tyrosine kinase p56lck in human Jurkat T cells.Biochem Pharmacol. 2011 Nov 1;82(9):1110-25. doi: 10.1016/j.bcp.2011.07.085. Epub 2011 Jul 23. Biochem Pharmacol. 2011. PMID: 21819973
-
The critical role of JNK in the ER-mitochondrial crosstalk during apoptotic cell death.J Cell Physiol. 2012 May;227(5):1791-5. doi: 10.1002/jcp.22903. J Cell Physiol. 2012. PMID: 21732347 Review.
Cited by
-
Viruses Run: The Evasion Mechanisms of the Antiviral Innate Immunity by Hantavirus.Front Microbiol. 2021 Sep 30;12:759198. doi: 10.3389/fmicb.2021.759198. eCollection 2021. Front Microbiol. 2021. PMID: 34659193 Free PMC article. Review.
-
Hantaviruses and TNF-alpha act synergistically to induce ERK1/2 inactivation in Vero E6 cells.Virol J. 2008 Sep 29;5:110. doi: 10.1186/1743-422X-5-110. Virol J. 2008. PMID: 18822184 Free PMC article.
-
The nucleocapsid protein of hantaviruses: much more than a genome-wrapping protein.Virus Genes. 2018 Feb;54(1):5-16. doi: 10.1007/s11262-017-1522-3. Epub 2017 Nov 20. Virus Genes. 2018. PMID: 29159494 Review.
-
Birth and pathogenesis of rogue respiratory viruses.Annu Rev Pathol. 2015;10:449-71. doi: 10.1146/annurev-pathol-012414-040501. Epub 2014 Nov 24. Annu Rev Pathol. 2015. PMID: 25423349 Free PMC article. Review.
-
Dengue virus serotype infection specifies the activation of the unfolded protein response.Virol J. 2007 Sep 24;4:91. doi: 10.1186/1743-422X-4-91. Virol J. 2007. PMID: 17888185 Free PMC article.
References
-
- Akhmatova N.K., Yusupova R.S., Khaiboullina S.F., Sibiryak S.V. Lymphocyte apoptosis during hemorrhagic fever with renal syndrome. Russ. J. Immunol. 2003;8:37–46. - PubMed
-
- Breckenridge D.G., Germain M., Mathai J.P., Nguyen M., Shore G.C. Regulation of apoptosis by endoplasmic reticulum pathways. Oncogene. 2003;22:8608–8618. - PubMed
-
- Choukhi A., Pillez A., Drobecq H., Sergheraert C., Wychowski C., Dubuisson J. Characterization of aggregates of hepatitis C virus glycoproteins. J. Gen. Virol. 1999;80:3099–3107. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
