

Lineage relationships, homeostasis, and recall capacities of central- and effector-memory CD8 T cells in vivo
- PMID: 15710650
- PMCID: PMC2213051
- DOI: 10.1084/jem.20040876
Lineage relationships, homeostasis, and recall capacities of central- and effector-memory CD8 T cells in vivo
Abstract
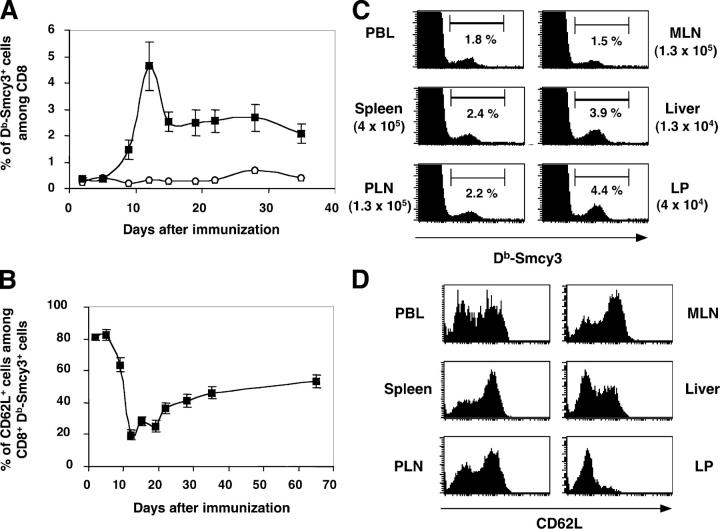
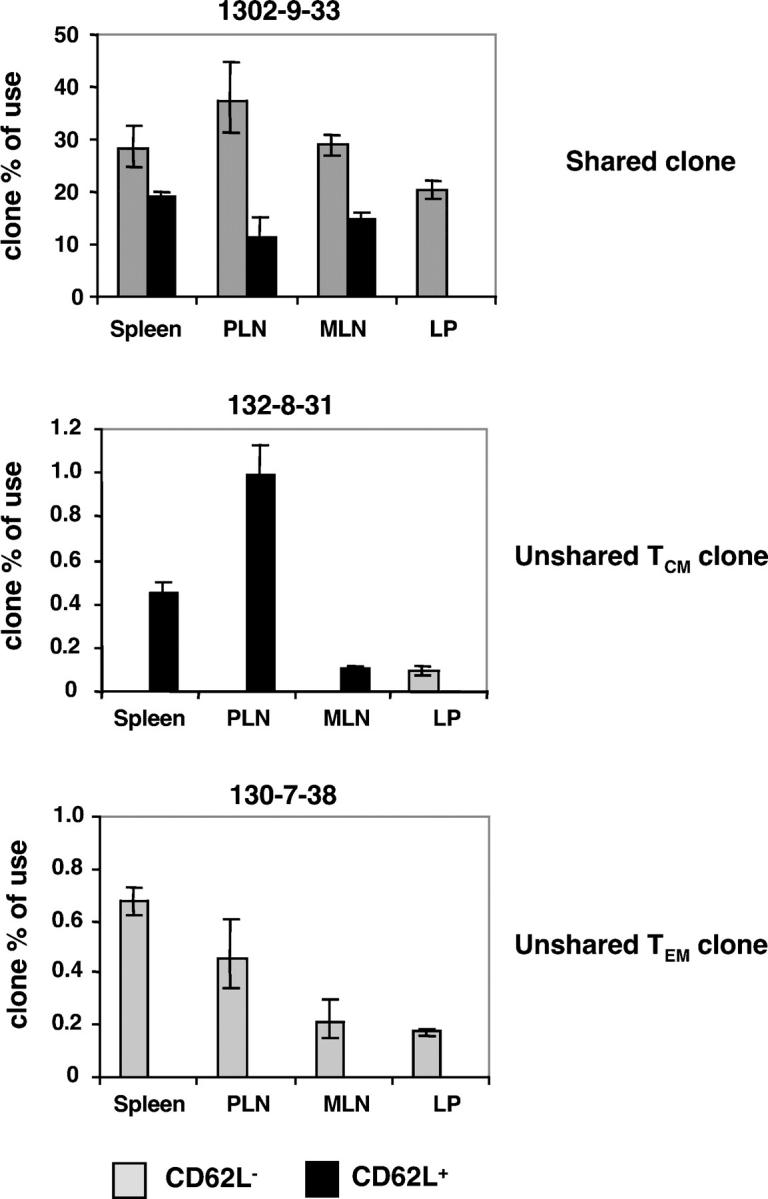
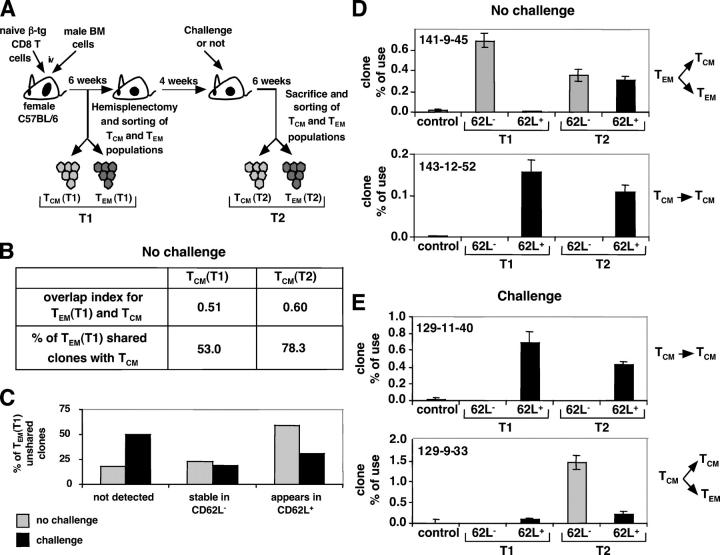
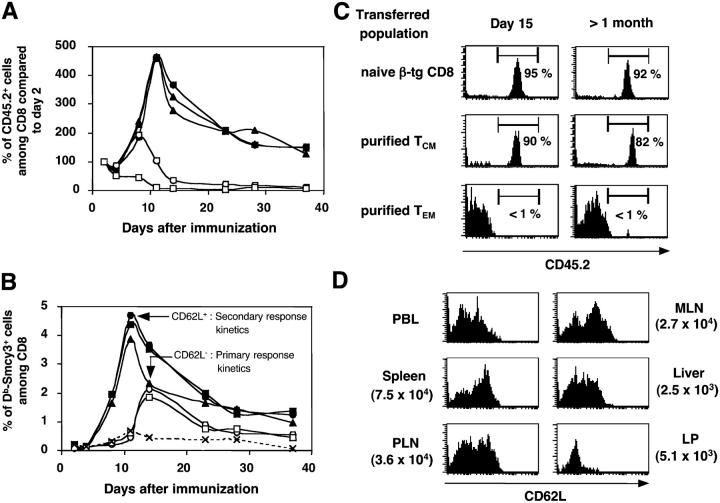
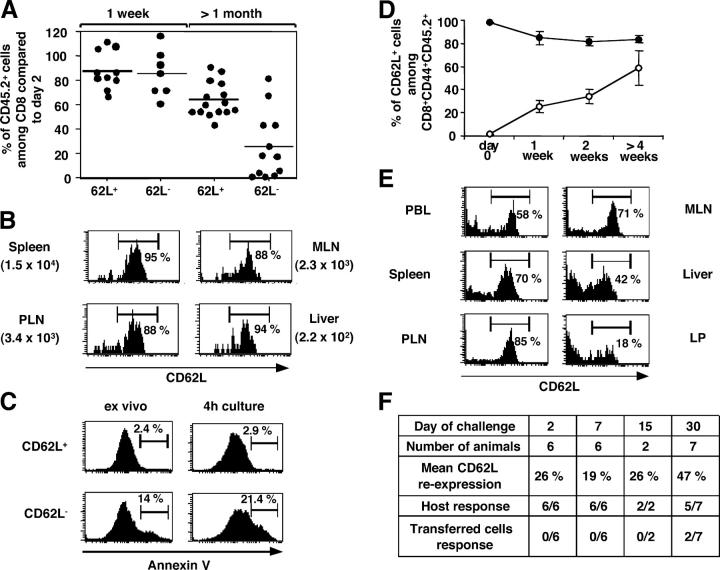
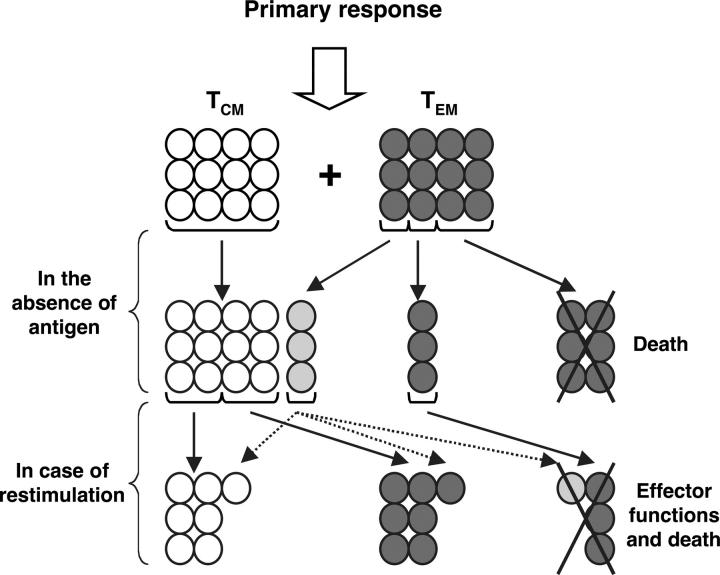
The lineage relationships of central-memory T cells (T(CM)) cells and effector-memory T cells (T(EM)), as well as their homeostasis and recall capacities, are still controversial. We investigated these issues in a murine model using two complementary approaches: T cell receptor repertoire analysis and adoptive transfer experiments of purified H-Y-specific T(CM) and T(EM) populations. Repertoire studies showed that approximately two thirds of T(CM) and T(EM) clones derived from a common naive precursor, whereas the other third was distinct. Both approaches highlighted that T(CM) and T(EM) had drastically distinct behaviors in vivo, both in the absence of antigen or upon restimulation. T(CM) clones were stable in the absence of restimulation and mounted a potent and sustained recall response upon secondary challenge, giving rise to both T(CM) and T(EM), although only a fraction of T(CM) generated T(EM). In contrast, T(EM) persisted for only a short time in the absence of antigen and, although a fraction of them were able to express CD62L, they were unable to mount a proliferative response upon secondary challenge in this model.
Figures
References
-
- Ahmed, R., and D. Gray. 1996. Immunological memory and protective immunity: understanding their relation. Science. 272:54–60. - PubMed
-
- McHeyzer-Williams, M.G., and R. Ahmed. 1999. B cell memory and the long-lived plasma cell. Curr. Opin. Immunol. 11:172–179. - PubMed
-
- Sprent, J., and C.D. Surh. 2002. T cell memory. Annu. Rev. Immunol. 20:551–579. - PubMed
-
- Woodland, D.L., and R.W. Dutton. 2003. Heterogeneity of CD4(+) and CD8(+) T cells. Curr. Opin. Immunol. 15:336–342. - PubMed
-
- Lefrancois, L., and D. Masopust. 2002. T cell immunity in lymphoid and non-lymphoid tissues. Curr. Opin. Immunol. 14:503–508. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
