

Honey bees navigate according to a map-like spatial memory
- PMID: 15710880
- PMCID: PMC549458
- DOI: 10.1073/pnas.0408550102
Honey bees navigate according to a map-like spatial memory
Abstract
By using harmonic radar, we report the complete flight paths of displaced bees. Test bees forage at a feeder or are recruited by a waggle dance indicating the feeder. The flights are recorded after the bees are captured when leaving the hive or the feeder and are released at an unexpected release site. A sequence of behavioral routines become apparent: (i) initial straight flights in which they fly the course that they were on when captured (foraging bees) or that they learned during dance communication (recruited bees); (ii) slow search flights with frequent changes of direction in which they attempt to "get their bearings"; and (iii) straight and rapid flights directed either to the hive or first to the feeding station and then to the hive. These straight homing flights start at locations all around the hive and at distances far out of the visual catchment area around the hive or the feeding station. Two essential criteria of a map-like spatial memory are met by these results: bees can set course at any arbitrary location in their familiar area, and they can choose between at least two goals. This finding suggests a rich, map-like organization of spatial memory in navigating honey bees.
Figures
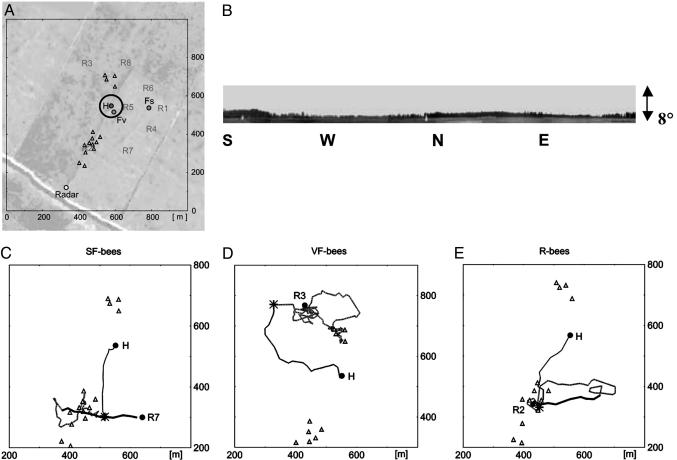
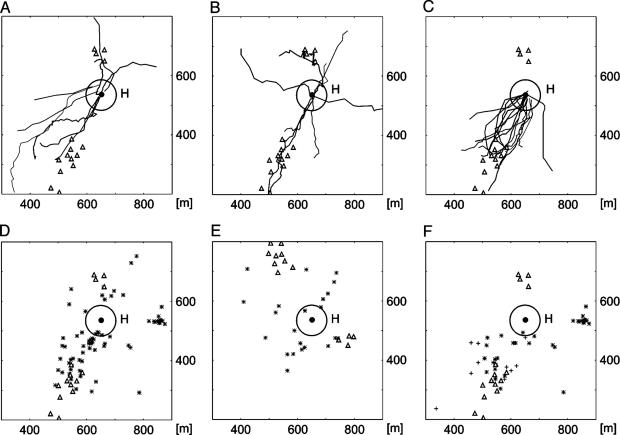
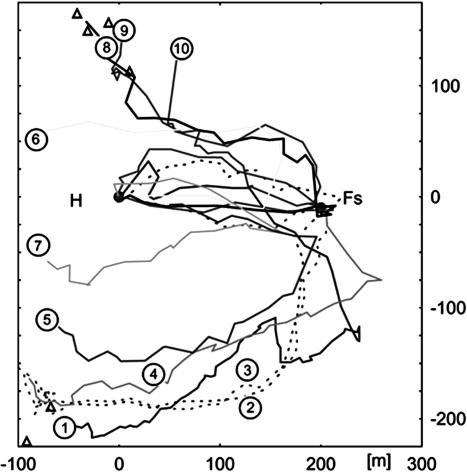
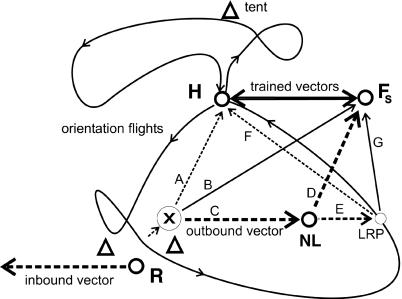
 ) at which a particular heading and distance are retrieved (A), they may either follow this information and return to the hive or switch their motivation and aim for the feeder. In such a situation they retrieve the vector components (heading and distance) of the outbound route flight from the hive-feeder to the feeder (C), which leads them to a new location (NL), or they perform some form of large-scale vector integration that leads them along a novel route to the feeder (B). At the novel location NL they may apply the same vector integration (D) or reach another known place (LRP) from which they go through the same procedure, use the associated flight parameters (F) to fly back to the hive directly or perform vector integration (G). However, our data also are consistent with the possibility that bees establish during orientation flights a relational map-like memory. In that case, they would localize themselves according to local landmarks and views, and they would choose a flight vector to the localizations of the hive or the feeder.
) at which a particular heading and distance are retrieved (A), they may either follow this information and return to the hive or switch their motivation and aim for the feeder. In such a situation they retrieve the vector components (heading and distance) of the outbound route flight from the hive-feeder to the feeder (C), which leads them to a new location (NL), or they perform some form of large-scale vector integration that leads them along a novel route to the feeder (B). At the novel location NL they may apply the same vector integration (D) or reach another known place (LRP) from which they go through the same procedure, use the associated flight parameters (F) to fly back to the hive directly or perform vector integration (G). However, our data also are consistent with the possibility that bees establish during orientation flights a relational map-like memory. In that case, they would localize themselves according to local landmarks and views, and they would choose a flight vector to the localizations of the hive or the feeder.References
-
- Wehner, R. (2003) J. Comp. Physiol. A 189, 579-588. - PubMed
-
- Gallistel, C. R. (1998) in Conceptual and Methodological Foundations—Vol. 4 of an Invitation to Cognitive Science, eds. Sternberg, S. & Scarborough, D. (MIT Press, Cambridge, MA), 2nd Ed., pp. 1-51.
-
- Collett, T. S. & Collett, M. (2002) Nat. Rev. Neurosci. 3, 542-552. - PubMed
-
- Wehner, R. & Menzel, R. (1990) Annu. Rev. Neurosci. 13, 403-414. - PubMed
-
- Mittelstaedt, H. (1983) in Fortschritte der Zoologie (Gustav Fischer, Stuttgart), pp. 197-212.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
