

The mutation Y1206S increases the affinity of the sulphonylurea receptor SUR2A for glibenclamide and enhances the effects of coexpression with Kir6.2
- PMID: 15711591
- PMCID: PMC1576091
- DOI: 10.1038/sj.bjp.0706142
The mutation Y1206S increases the affinity of the sulphonylurea receptor SUR2A for glibenclamide and enhances the effects of coexpression with Kir6.2
Abstract
1. ATP-sensitive K(+) channels (K(ATP) channels) are tetradimeric complexes of inwardly rectifying K(+) channels (Kir6.x) and sulphonylurea receptors (SURs). The SURs SUR2A (cardiac) and SUR2B (smooth muscle) differ only in the last 42 amino acids. In SUR2B, the mutation Y1206S, located at intracellular loop 8, increases the affinity for glibenclamide (GBC) about 10-fold. Here, we examined whether the mutation Y1206S in SUR2A had effects similar to those in SUR2B.2. GBC bound to SUR2A with K(D)=20 nM; the mutation increased affinity approximately 5 x. 3. In cells, coexpression of SUR2A with Kir6.2 increased the affinity for GBC approximately 3 x; with the mutant, the increase was 9 x. 4. The mutation did not affect the affinity of SUR2A for openers; coexpression with Kir6.2 reduced opener affinity of wild-type and mutant SUR2A by about 2 x. 5. The negative allosteric interaction between the opener, P1075, and GBC at wild-type and mutant SUR2A was markedly affected by the presence of MgATP and by coexpression with Kir6.2. 6. In inside-out patches, GBC inhibited the wild-type Kir6.2/SUR2A and 2B channels with IC(50) values of 27 nM; the mutation shifted the IC(50) values to approximately 1 nM. 7. The data show that the mutation Y1206S increased the affinity of SUR2A for GBC and modulated the effects of coexpression. Overall, the changes were similar to those observed with SUR2B(Y1206S), suggesting that the differences in the last 42 carboxy-terminal amino acids of SUR2A and 2B are of limited influence on the binding of GBC and P1075 to the SUR2 isoforms.
Figures
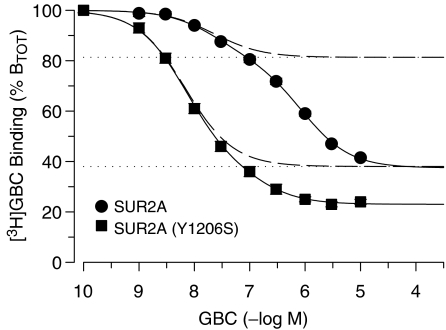
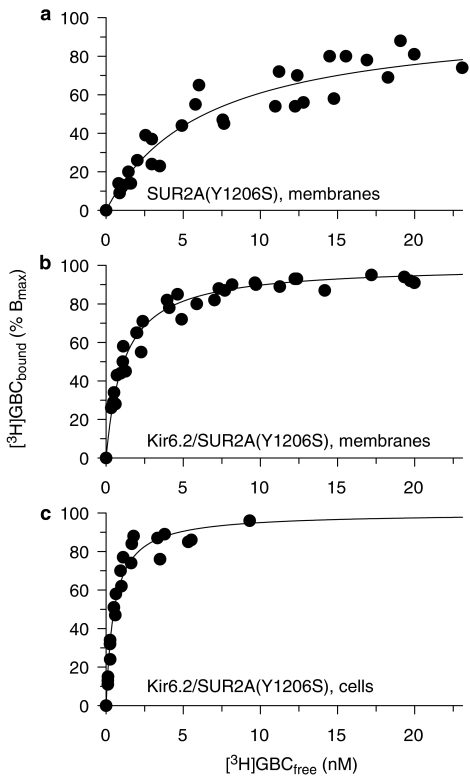
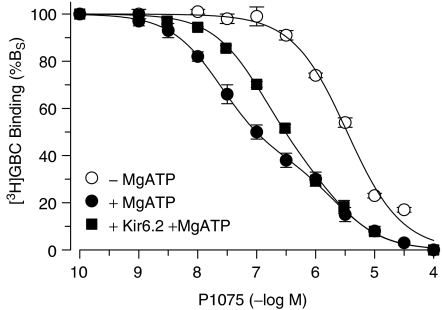
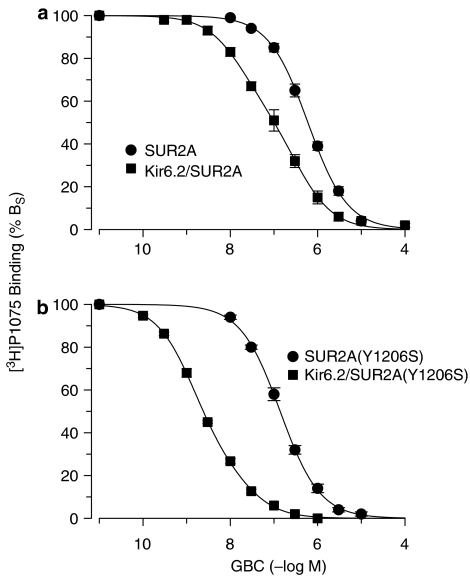
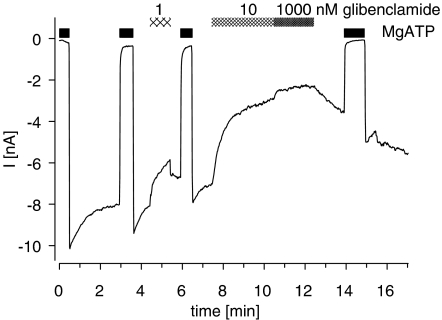
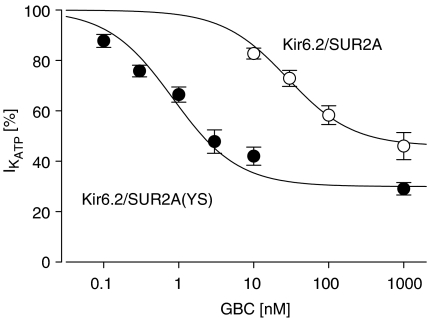
 ) prior to the application of GBC. Concentration dependencies were analysed using the Hill equation with nH=1; the parameters are listed in Table 4. n=4–10 per data point; data for wild type are from Russ et al. (2001).
) prior to the application of GBC. Concentration dependencies were analysed using the Hill equation with nH=1; the parameters are listed in Table 4. n=4–10 per data point; data for wild type are from Russ et al. (2001).References
-
- AGUILAR-BRYAN L., BRYAN J. Molecular biology of adenosine triphosphate-sensitive potassium channels. Endocrine Rev. 1999;20:101–135. - PubMed
-
- AGUILAR-BRYAN L., NICHOLS C.G., WECHSLER S.W., CLEMENT IV J.P., BOYD A.E., III, GONZÁLES G., HERRERA-SOZA H., NGUY K., BRYAN J., NELSON D.A. Cloning of the β cell high-affinity sulfonylurea receptor: a regulator of insulin secretion. Science. 1995;268:423–426. - PubMed
-
- ASHCROFT F.M., GRIBBLE F.M. Correlating structure and function in ATP-sensitive K+ channels. Trends Neurosci. 1998;21:288–294. - PubMed
-
- ASHCROFT S.J.H., ASHCROFT F.M. Properties and functions of ATP-sensitive K-channels. Cell. Signal. 1990;2:197–214. - PubMed
-
- ASHFIELD R., GRIBBLE F.M., ASHCROFT S.J.H., ASHCROFT F.M. Identification of the high-affinity tolbutamide site on the SUR1 subunit of the KATP channel. Diabetes. 1999;48:1341–1347. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
