

Mitogen-activated protein kinases with distinct requirements for Ste5 scaffolding influence signaling specificity in Saccharomyces cerevisiae
- PMID: 15713635
- PMCID: PMC549360
- DOI: 10.1128/MCB.25.5.1793-1803.2005
Mitogen-activated protein kinases with distinct requirements for Ste5 scaffolding influence signaling specificity in Saccharomyces cerevisiae
Abstract
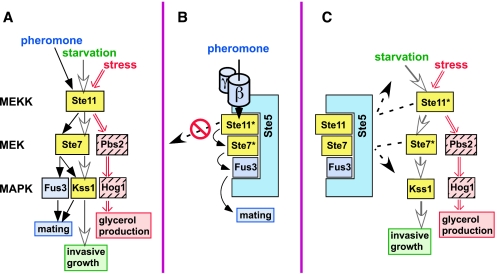
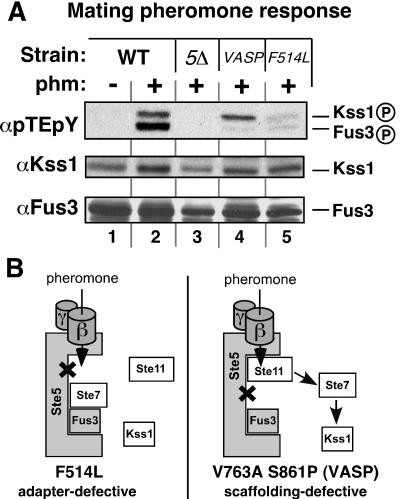
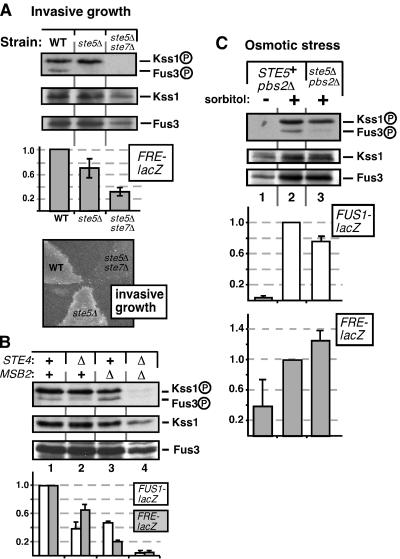
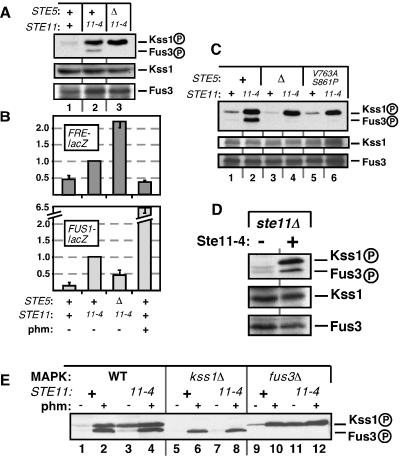
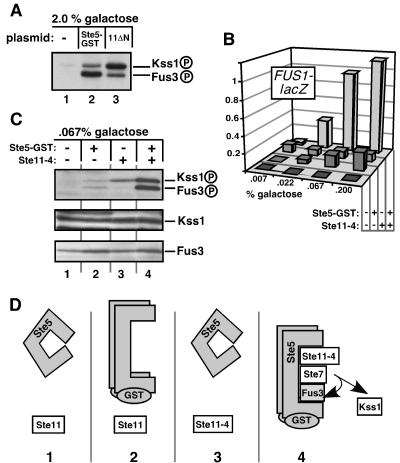
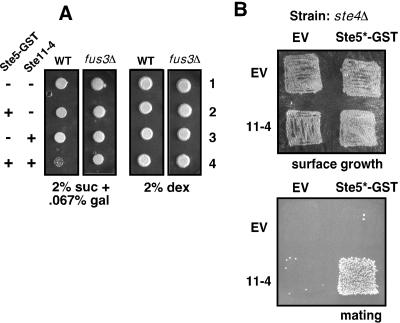
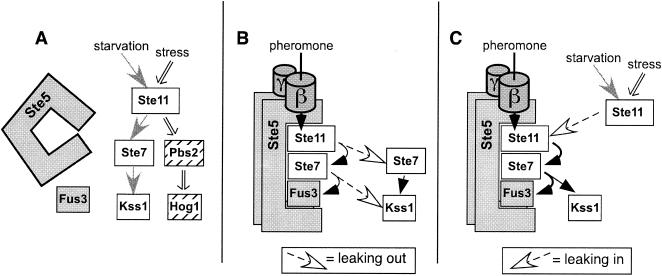
Scaffold proteins are believed to enhance specificity in cell signaling when different pathways share common components. The prototype scaffold Ste5 binds to multiple components of the Saccharomyces cerevisiae mating pheromone response pathway, thereby conducting the mating signal to the Fus3 mitogen-activated protein kinase (MAPK). Some of the kinases that Ste5 binds to, however, are also shared with other pathways. Thus, it has been presumed that Ste5 prevents its bound kinases from transgressing into other pathways and protects them from intrusions from those pathways. Here we found that Fus3MAPK required Ste5 scaffolding to receive legitimate signals from the mating pathway as well as misdirected signals leaking from other pathways. Furthermore, increasing the cellular concentration of active Ste5 enhanced the channeling of inappropriate stimuli to Fus3. This aberrant signal crossover resulted in the erroneous induction of cell cycle arrest and mating. In contrast to Fus3, the Kss1 MAPK did not require Ste5 scaffolding to receive either authentic or leaking signals. Furthermore, the Ste11 kinase, once activated via Ste5, was able to signal to Kss1 independently of Ste5 scaffolding. These results argue that Ste5 does not act as a barrier that actively prevents signal crossover to Fus3 and that Ste5 may not effectively sequester its activated kinases away from other pathways. Rather, we suggest that specificity in this network is promoted by the selective activation of Ste5 and the distinct requirements of the MAPKs for Ste5 scaffolding.
Figures
References
-
- Bardwell, L., J. G. Cook, E. C. Chang, B. R. Cairns, and J. Thorner. 1996. Signaling in the yeast pheromone response pathway: specific and high-affinity interaction of the mitogen-activated protein (MAP) kinases Kss1 and Fus3 with the upstream MAP kinase kinase Ste7. Mol. Cell. Biol. 16:3637-3650. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases