

Formation of an active tissue-specific chromatin domain initiated by epigenetic marking at the embryonic stem cell stage
- PMID: 15713636
- PMCID: PMC549375
- DOI: 10.1128/MCB.25.5.1804-1820.2005
Formation of an active tissue-specific chromatin domain initiated by epigenetic marking at the embryonic stem cell stage
Abstract
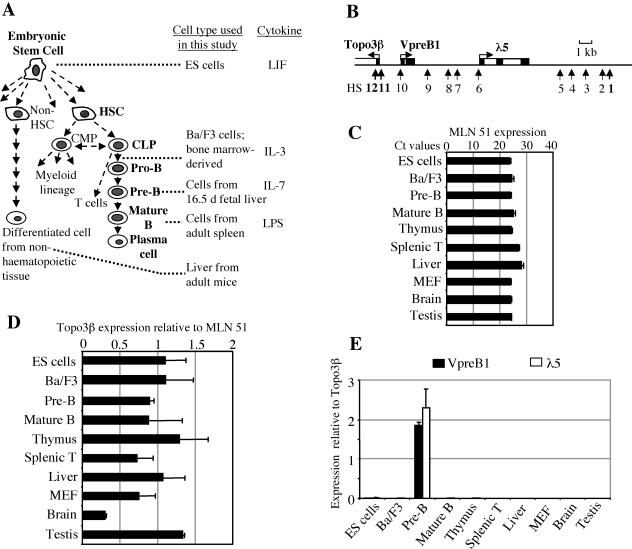
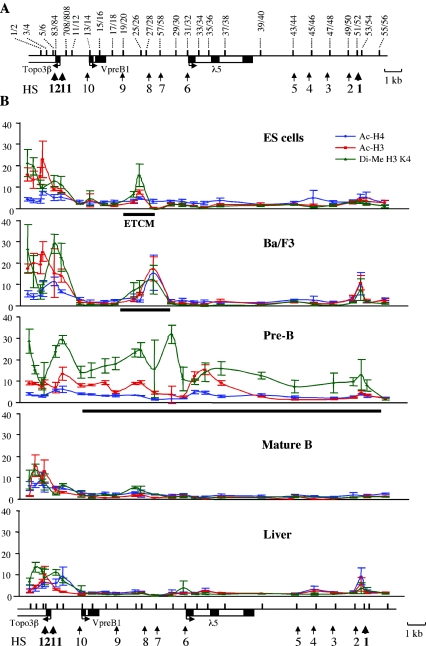
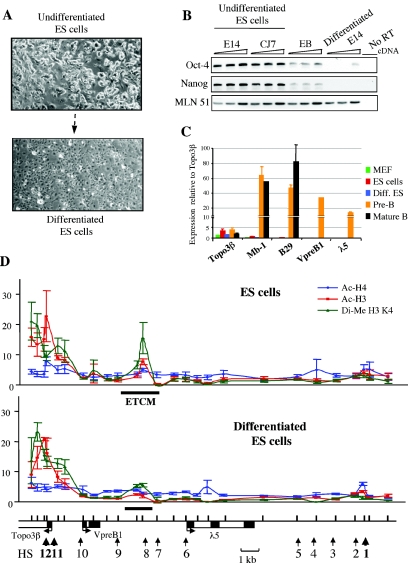
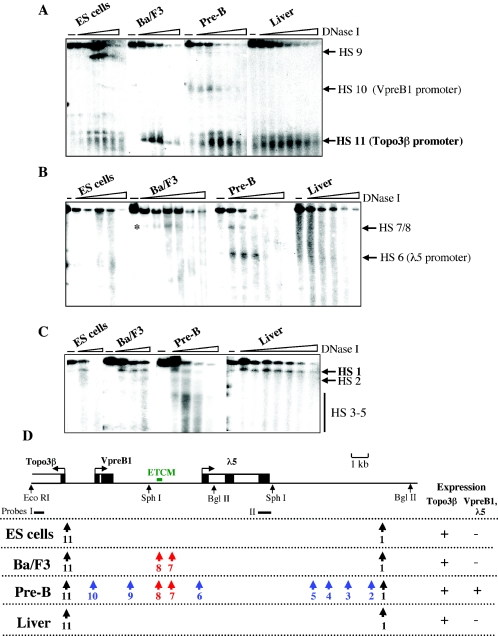
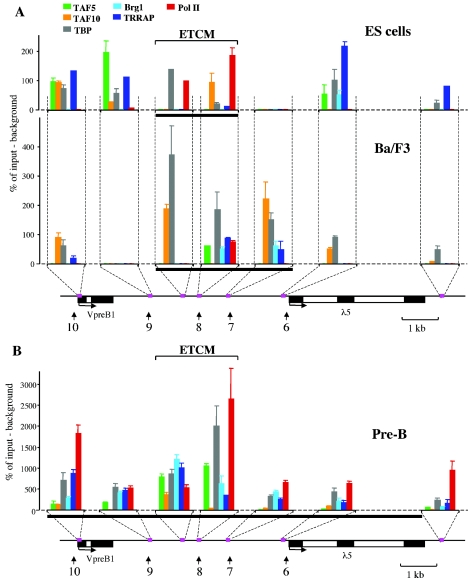
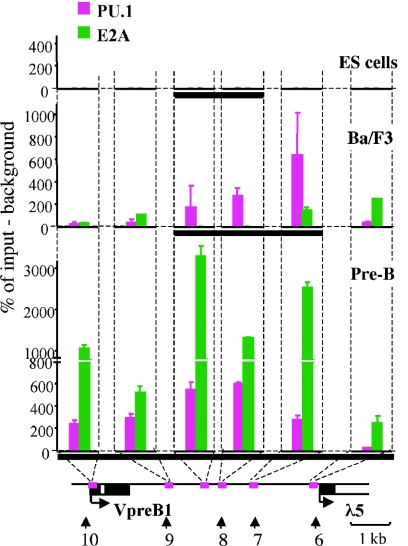
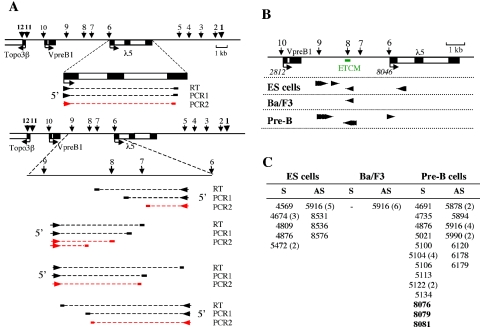
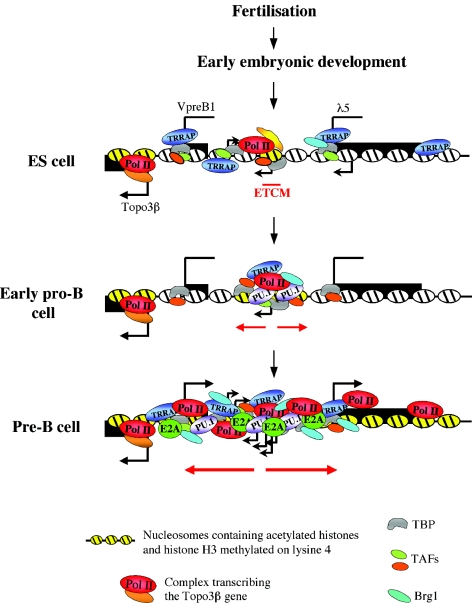
The differentiation potential of stem cells is determined by the ability of these cells to establish and maintain developmentally regulated gene expression programs that are specific to different lineages. Although transcriptionally potentiated epigenetic states of genes have been described for haematopoietic progenitors, the developmental stage at which the formation of lineage-specific gene expression domains is initiated remains unclear. In this study, we show that an intergenic cis-acting element in the mouse lambda5-VpreB1 locus is marked by histone H3 acetylation and histone H3 lysine 4 methylation at a discrete site in embryonic stem (ES) cells. The epigenetic modifications spread from this site toward the VpreB1 and lambda5 genes at later stages of B-cell development, and a large, active chromatin domain is established in pre-B cells when the genes are fully expressed. In early B-cell progenitors, the binding of haematopoietic factor PU.1 coincides with the expansion of the marked region, and the region becomes a center for the recruitment of general transcription factors and RNA polymerase II. In pre-B cells, E2A also binds to the locus, and general transcription factors are distributed across the active domain, including the gene promoters and the intergenic region. These results suggest that localized epigenetic marking is important for establishing the transcriptional competence of the lambda5 and VpreB1 genes as early as the pluripotent ES cell stage.
Figures
References
-
- Akashi, K., D. Traver, T. Miyamoto, and I. L. Weissman. 2000. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature 404:193-197. - PubMed
-
- Anderson, M. K., G. Hernandez-Hoyos, R. A. Diamond, and E. V. Rothenberg. 1999. Precise developmental regulation of Ets family transcription factors during specification and commitment to the T cell lineage. Development 126:3131-3148. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases