

Structure of a mitochondrial supercomplex formed by respiratory-chain complexes I and III
- PMID: 15713802
- PMCID: PMC552927
- DOI: 10.1073/pnas.0408870102
Structure of a mitochondrial supercomplex formed by respiratory-chain complexes I and III
Abstract
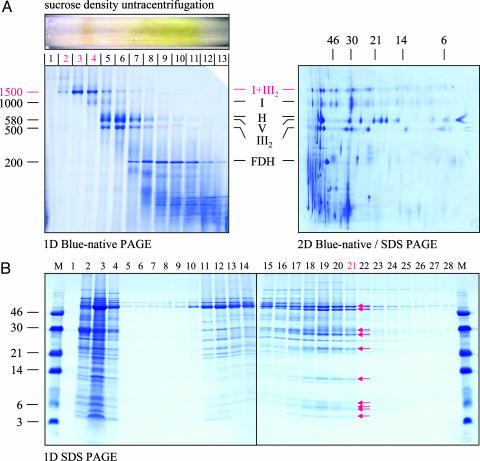
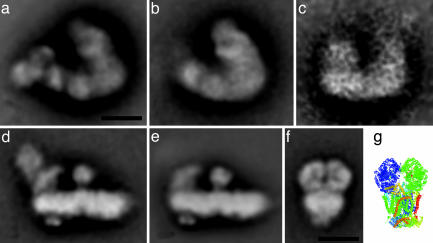
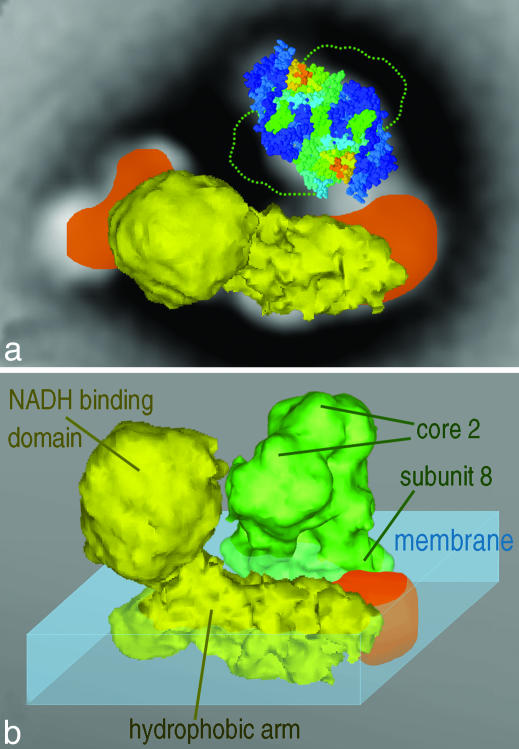
Mitochondria are central to the efficient provision of energy for eukaryotic cells. The oxidative-phosphorylation system of mitochondria consists of a series of five major membrane complexes: NADH-ubiquinone oxidoreductase (commonly known as complex I), succinate-ubiquinone oxidoreductase (complex II), ubiquinol-cytochrome c oxidoreductase (cytochrome bc1 complex or complex III), cytochrome c-O2 oxidoreductase (complex IV), and F1F0-ATP synthase (complex V). Several lines of evidence have recently suggested that complexes I and III-V might interact to form supercomplexes. However, because of their fragility, the structures of these supercomplexes are still unknown. A stable supercomplex consisting of complex I and dimeric complex III was purified from plant mitochondria. Structural characterization by single-particle EM indicates a specific type of interaction between monomeric complex I and dimeric complex III in a 1:1 ratio. We present a model for how complexes I and III are spatially organized within the I+III2 supercomplex.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
