

Gene expression analysis of the function of the male-specific lethal complex in Drosophila
- PMID: 15716510
- PMCID: PMC1449592
- DOI: 10.1534/genetics.104.036020
Gene expression analysis of the function of the male-specific lethal complex in Drosophila
Abstract
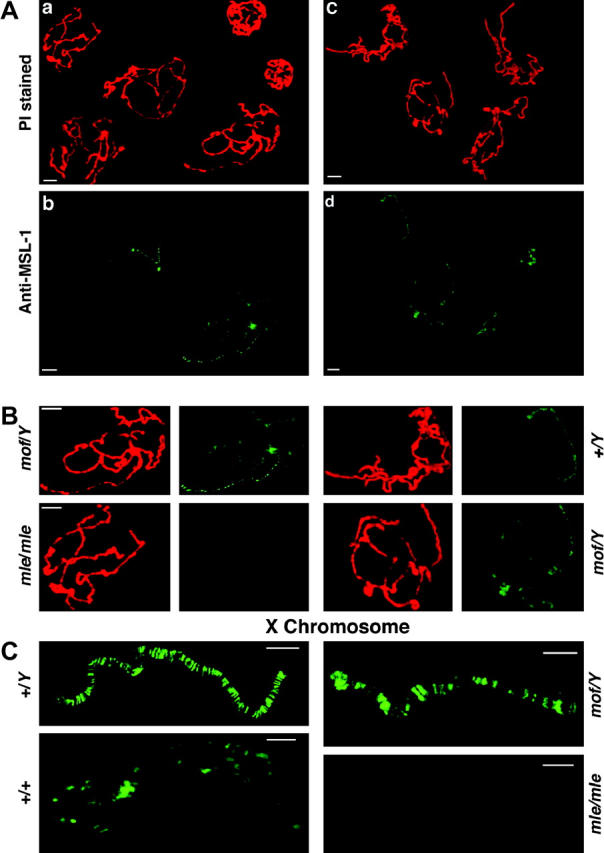
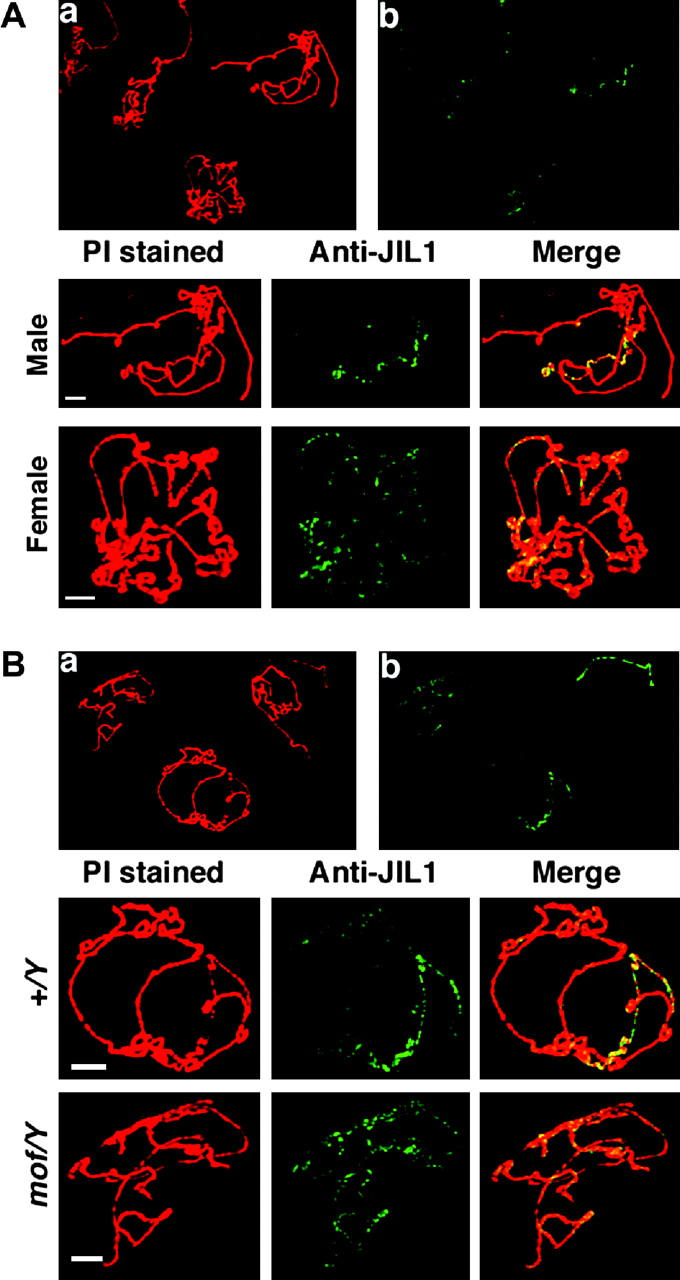
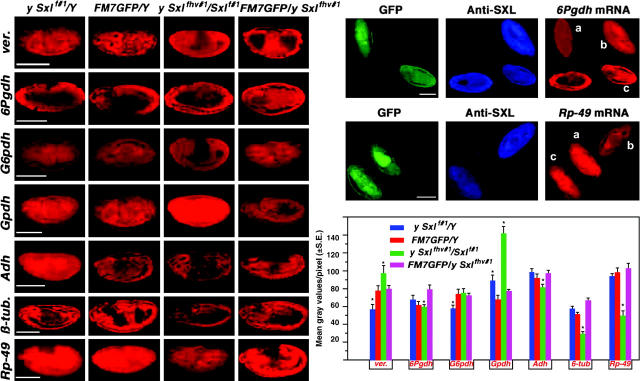
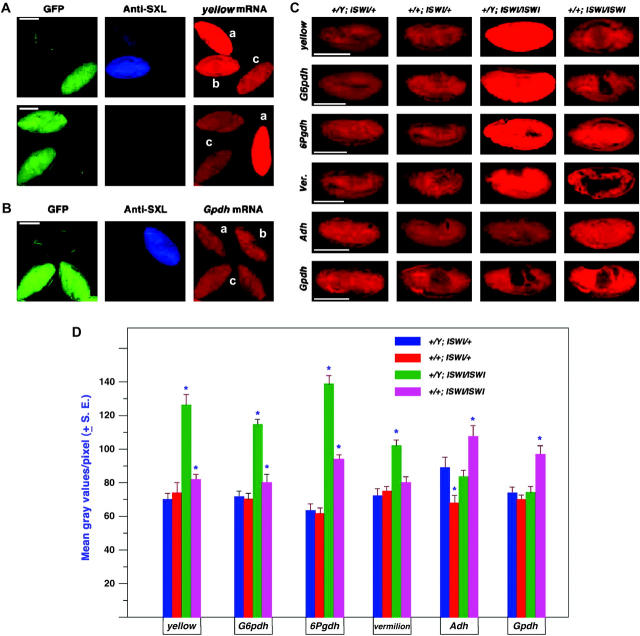
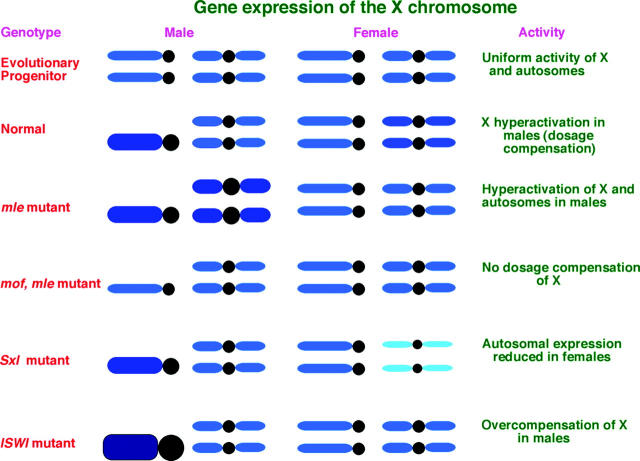
Dosage compensation refers to the equal expression of X-linked genes despite the difference in copy number between the two sexes. The male-specific lethal (MSL) complex is concentrated on the X chromosome in males. A gene expression assay for embryos was developed to examine the function of this complex. In mutant male embryos without either the MSL complex or MOF histone acetylase, dosage compensation is retained but autosomal expression is increased. Dosage compensation is lost in the double-mutant embryos. In embryos in which the MSL complex and MOF are targeted to the X chromosomes in females, the results are consistent with previous surveys showing that in general the X expression remains unchanged, but autosomal expression is reduced. Mutations in the ISWI chromatin-remodeling component cause increases specifically of X-linked genes in males. Thus, the function of the MSL complex in conjunction with ISWI is postulated to override the effect on gene expression of high histone acetylation on the male X. The basic determinant of dosage compensation is suggested to be an inverse dosage effect produced by an imbalance of transcription factors on the X vs. the autosomes. The sequestration of the MSL complex to the male X may have evolved to counteract a similar effect on the autosomes and to prevent an overexpression of the X chromosome in males that would otherwise occur due to the high levels of histone acetylation.
Figures
Similar articles
-
Dosage dependent gene regulation and the compensation of the X chromosome in Drosophila males.Genetica. 2003 Mar;117(2-3):179-90. doi: 10.1023/a:1022935927763. Genetica. 2003. PMID: 12723697 Review.
-
Role of the male specific lethal (msl) genes in modifying the effects of sex chromosomal dosage in Drosophila.Genetics. 1999 May;152(1):249-68. doi: 10.1093/genetics/152.1.249. Genetics. 1999. PMID: 10224258 Free PMC article.
-
Interaction study of the male specific lethal (MSL) complex and trans-acting dosage effects in metafemales of Drosophila melanogaster.Cytogenet Genome Res. 2009;124(3-4):298-311. doi: 10.1159/000218134. Epub 2009 Jun 25. Cytogenet Genome Res. 2009. PMID: 19556782 Free PMC article.
-
Evidence that MSL-mediated dosage compensation in Drosophila begins at blastoderm.Development. 1996 Sep;122(9):2751-60. doi: 10.1242/dev.122.9.2751. Development. 1996. PMID: 8787749
-
The right dose for every sex.Chromosoma. 2007 Apr;116(2):95-106. doi: 10.1007/s00412-006-0089-x. Epub 2006 Nov 24. Chromosoma. 2007. PMID: 17124606 Free PMC article. Review.
Cited by
-
X-chromosome hyperactivation in mammals via nonlinear relationships between chromatin states and transcription.Nat Struct Mol Biol. 2011 Dec 4;19(1):56-61. doi: 10.1038/nsmb.2195. Nat Struct Mol Biol. 2011. PMID: 22139016 Free PMC article.
-
Studies on the short range spreading of the male specific lethal (MSL) complex on the X chromosome in Drosophila.Cytogenet Genome Res. 2009;124(2):158-69. doi: 10.1159/000207524. Epub 2009 May 5. Cytogenet Genome Res. 2009. PMID: 19420929 Free PMC article.
-
Male X-linked genes in Drosophila melanogaster are compensated independently of the Male-Specific Lethal complex.Epigenetics Chromatin. 2013 Oct 26;6(1):35. doi: 10.1186/1756-8935-6-35. Epigenetics Chromatin. 2013. PMID: 24279328 Free PMC article.
-
Regulatory RNAs and chromatin modification in dosage compensation: a continuous path from flies to humans?Reprod Biol Endocrinol. 2008 Mar 20;6:12. doi: 10.1186/1477-7827-6-12. Reprod Biol Endocrinol. 2008. PMID: 18355403 Free PMC article. Review.
-
Dosage compensation, the origin and the afterlife of sex chromosomes.Chromosome Res. 2006;14(4):417-31. doi: 10.1007/s10577-006-1064-3. Chromosome Res. 2006. PMID: 16821137 Review.
References
-
- Akhtar, A., and P. B. Becker, 2000. Activation of transcription through histone H4 acetylation by MOF, an acetyltransferase essential for dosage compensation in Drosophila. Mol. Cell 5: 367–375. - PubMed
-
- Bahn, S., M. Mimmack, M. Ryan, M. A. Caldwell, E. Jauniaux et al., 2002. Neuronal target genes of the neuron-restrictive silencer factor in neurospheres derived from fetuses with Down's syndrome: a gene expression study. Lancet 359: 310–315. - PubMed
-
- Belote, J. M., and J. C. Lucchesi, 1980. Control of X chromosome transcription by the maleless gene in Drosophila. Nature 285: 573–575. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
