

Modulation of CD1d-restricted NKT cell responses by using N-acyl variants of alpha-galactosylceramides
- PMID: 15722411
- PMCID: PMC552918
- DOI: 10.1073/pnas.0407488102
Modulation of CD1d-restricted NKT cell responses by using N-acyl variants of alpha-galactosylceramides
Abstract
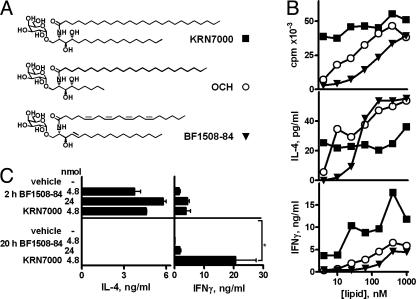
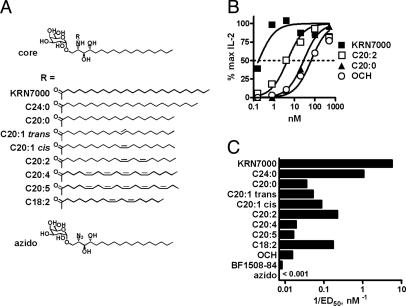
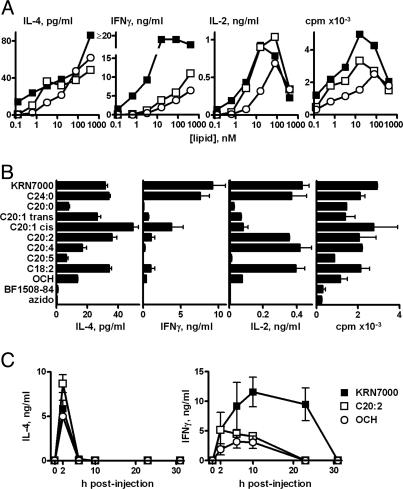
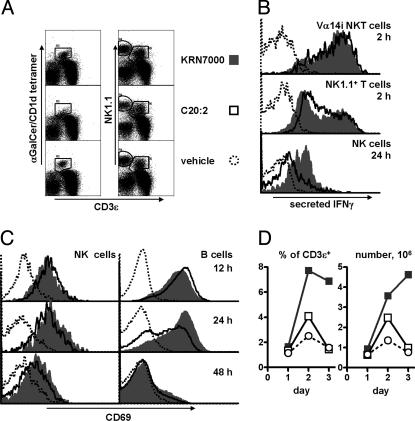
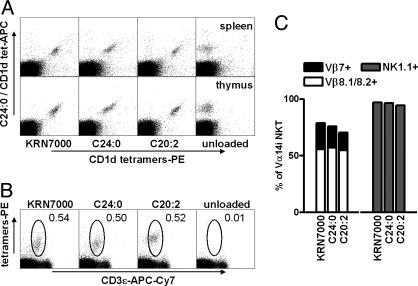
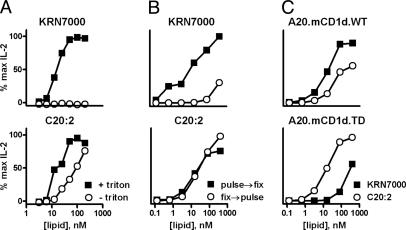
A form of alpha-galactosylceramide, KRN7000, activates CD1d-restricted Valpha14-invariant (Valpha14i) natural killer (NK) T cells and initiates multiple downstream immune reactions. We report that substituting the C26:0 N-acyl chain of KRN7000 with shorter, unsaturated fatty acids modifies the outcome of Valpha14i NKT cell activation. One analogue containing a diunsaturated C20 fatty acid (C20:2) potently induced a T helper type 2-biased cytokine response, with diminished IFN-gamma production and reduced Valpha14i NKT cell expansion. C20:2 also exhibited less stringent requirements for loading onto CD1d than KRN7000, suggesting a mechanism for the immunomodulatory properties of this lipid. The differential cellular response elicited by this class of Valpha14i NKT cell agonists may prove to be useful in immunotherapeutic applications.
Figures
References
-
- Brossay, L. & Kronenberg, M. (1999) Immunogenetics 50, 146-151. - PubMed
-
- Godfrey, D. I., Hammond, K. J., Poulton, L. D., Smyth, M. J. & Baxter, A. G. (2000) Immunol. Today 21, 573-583. - PubMed
-
- Wilson, S. B. & Delovitch, T. L. (2003) Nat. Rev. Immunol. 3, 211-222. - PubMed
-
- Poulton, L. D., Smyth, M. J., Hawke, C. G., Silveira, P., Shepherd, D., Naidenko, O. V., Godfrey, D. I. & Baxter, A. G. (2001) Int. Immunol. 13, 887-896. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
