

A molecular-mechanical model of the microtubule
- PMID: 15722432
- PMCID: PMC1305467
- DOI: 10.1529/biophysj.104.051789
A molecular-mechanical model of the microtubule
Abstract
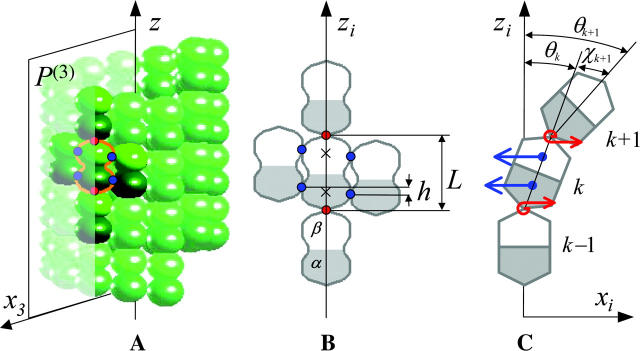
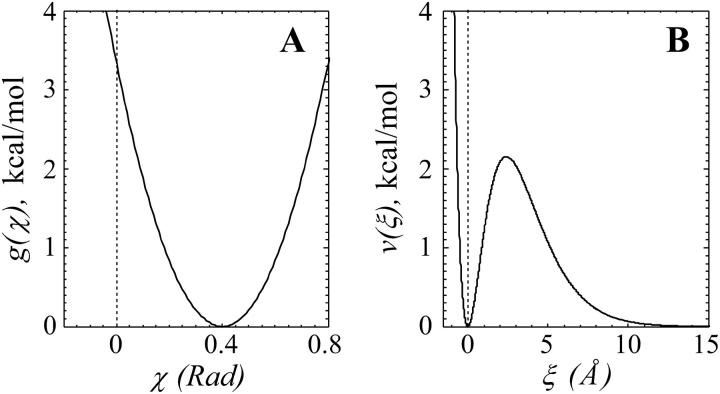
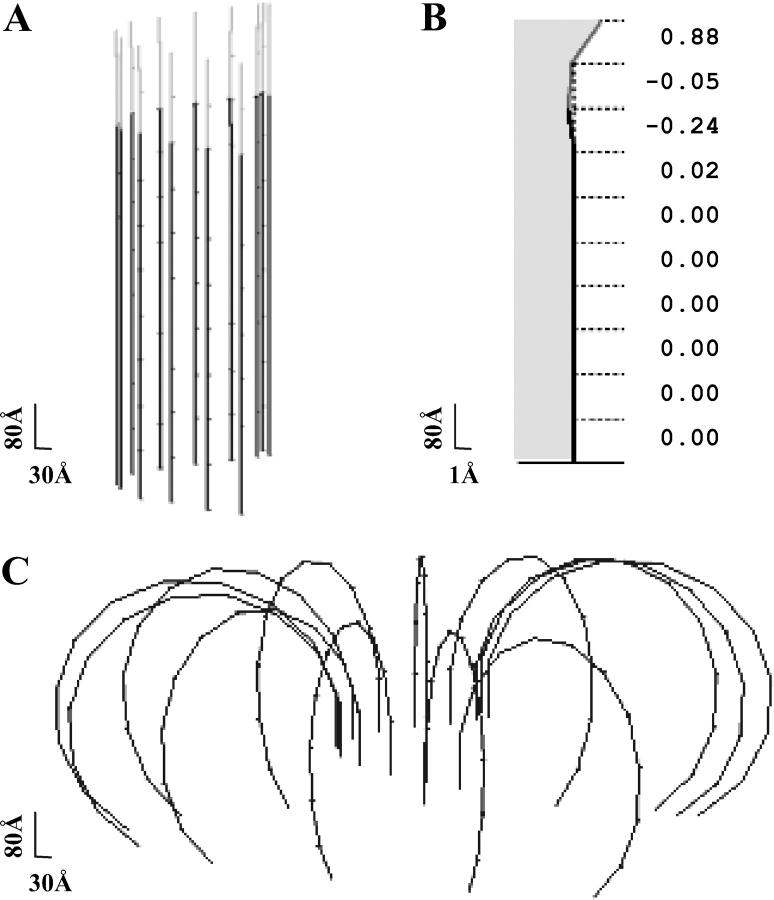
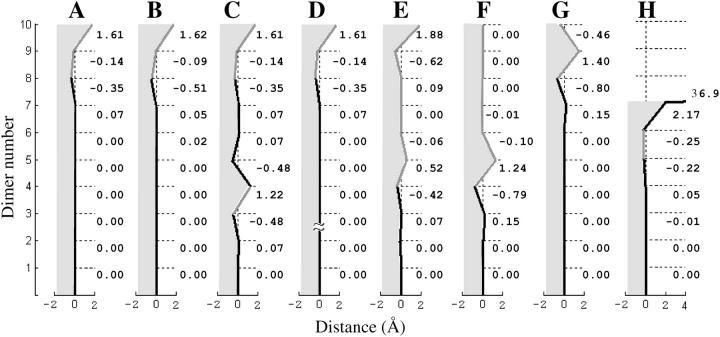
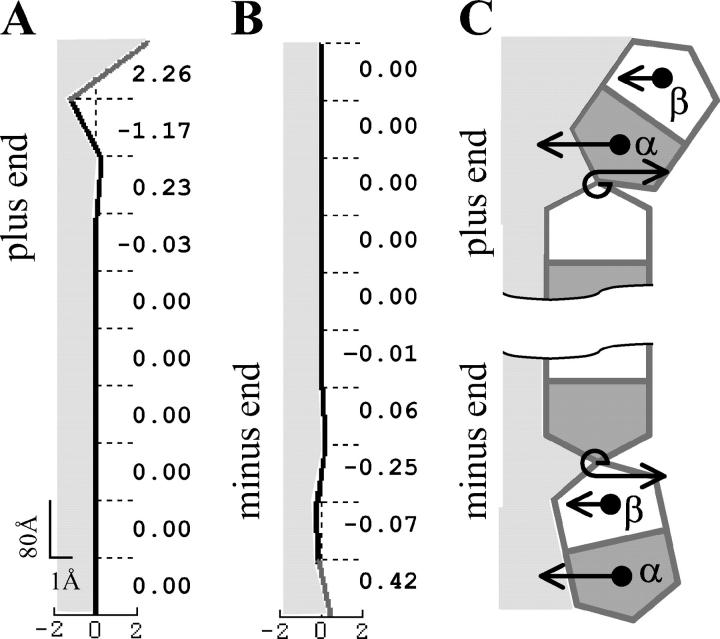
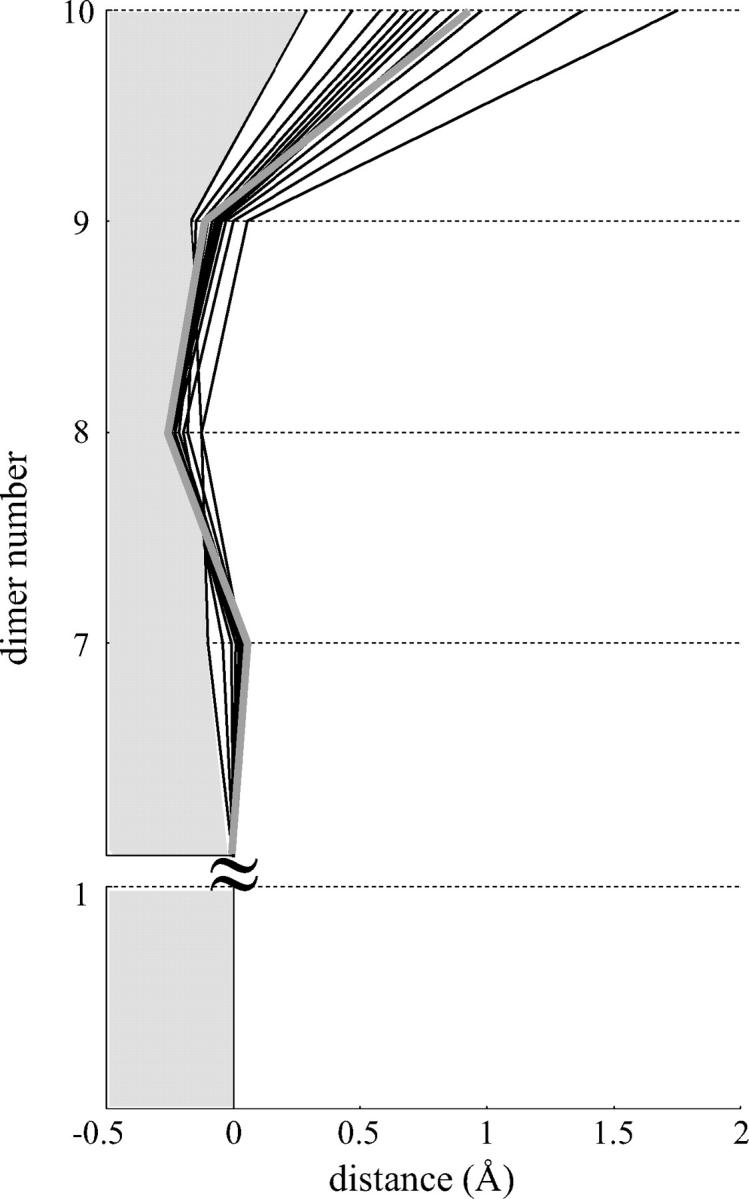
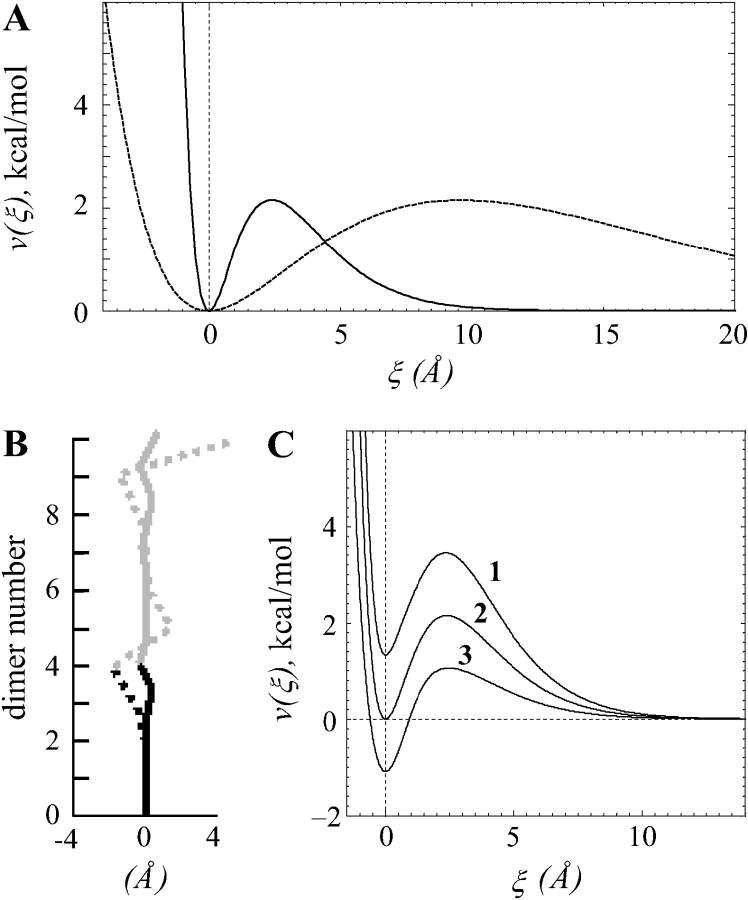
Dynamic instability of MTs is thought to be regulated by biochemical transformations within tubulin dimers that are coupled to the hydrolysis of bound GTP. Structural studies of nucleotide-bound tubulin dimers have recently provided a concrete basis for understanding how these transformations may contribute to MT dynamic instability. To analyze these ideas, we have developed a molecular-mechanical model in which structural and biochemical properties of tubulin are used to predict the shape and stability of MTs. From simple and explicit features of tubulin, we define bond energy relationships and explore the impact of their variations on integral MT properties. This modeling provides quantitative predictions about the GTP cap. It specifies important mechanical features underlying MT instability and shows that this property does not require GTP-hydrolysis to alter the strength of tubulin-tubulin bonds. The MT plus end is stabilized by at least two layers of GTP-tubulin subunits, whereas the minus end requires at least one; this and other differences between the ends are explained by asymmetric force balances. Overall, this model provides a new link between the biophysical characteristics of tubulin and the physiological behavior of MTs. It will also be useful in building a more complete description of MT dynamics and mechanics.
Figures
References
-
- Bayley, P., M. Schilstra, and S. Martin. 1989. A lateral cap model of microtubule dynamic instability. FEBS Lett. 259:181–184. - PubMed
-
- Caplow, M. 1992. Microtubule dynamics. Curr. Opin. Cell Biol. 4:58–65. - PubMed
-
- Caplow, M., and L. Fee. 2003. Concerning the chemical nature of tubulin subunits that cap and stabilize microtubules. Biochemistry. 42:2122–2126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
