

Structural basis for the interaction between pectin methylesterase and a specific inhibitor protein
- PMID: 15722470
- PMCID: PMC1069703
- DOI: 10.1105/tpc.104.028886
Structural basis for the interaction between pectin methylesterase and a specific inhibitor protein
Abstract
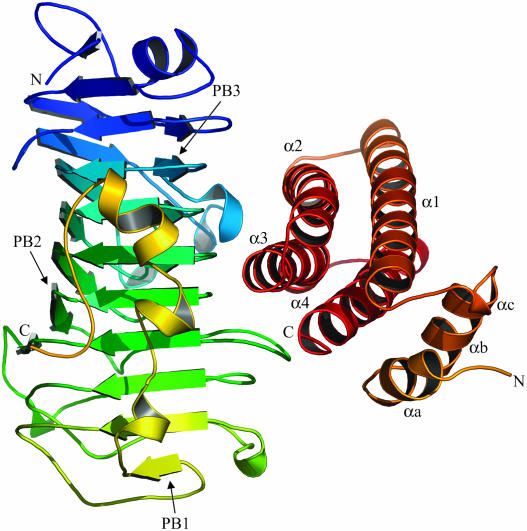
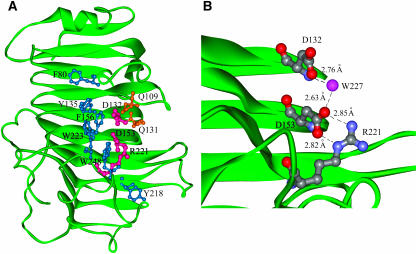
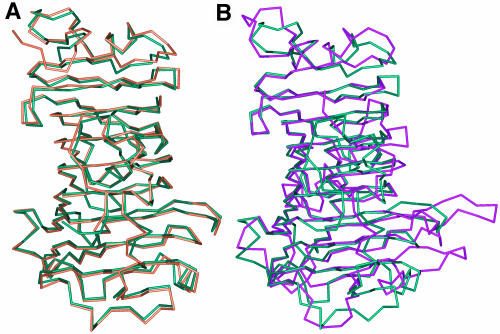
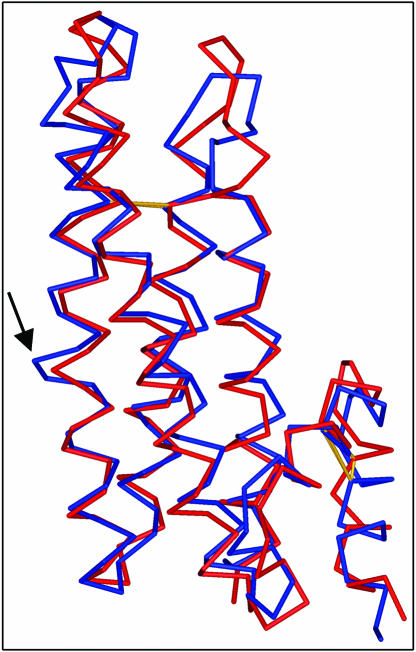
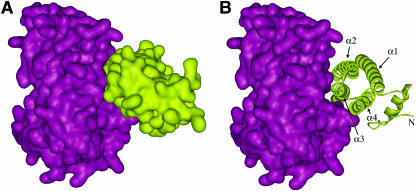
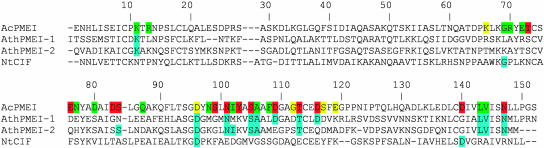
Pectin, one of the main components of the plant cell wall, is secreted in a highly methyl-esterified form and subsequently deesterified in muro by pectin methylesterases (PMEs). In many developmental processes, PMEs are regulated by either differential expression or posttranslational control by protein inhibitors (PMEIs). PMEIs are typically active against plant PMEs and ineffective against microbial enzymes. Here, we describe the three-dimensional structure of the complex between the most abundant PME isoform from tomato fruit (Lycopersicon esculentum) and PMEI from kiwi (Actinidia deliciosa) at 1.9-A resolution. The enzyme folds into a right-handed parallel beta-helical structure typical of pectic enzymes. The inhibitor is almost all helical, with four long alpha-helices aligned in an antiparallel manner in a classical up-and-down four-helical bundle. The two proteins form a stoichiometric 1:1 complex in which the inhibitor covers the shallow cleft of the enzyme where the putative active site is located. The four-helix bundle of the inhibitor packs roughly perpendicular to the main axis of the parallel beta-helix of PME, and three helices of the bundle interact with the enzyme. The interaction interface displays a polar character, typical of nonobligate complexes formed by soluble proteins. The structure of the complex gives an insight into the specificity of the inhibitor toward plant PMEs and the mechanism of regulation of these enzymes.
Figures

References
-
- Balestrieri, C., Castaldo, D., Giovane, A., Quagliuolo, L., and Servillo, L. (1990). A glycoprotein inhibitor of pectin methylesterase in kiwi fruit (Actinidia chinensis). Eur. J. Biochem. 193, 183–187. - PubMed
-
- Camardella, L., Carratore, V., Ciardiello, M.A., Servillo, L., Balestrieri, C., and Giovane, A. (2000). Kiwi protein inhibitor of pectin methylesterase amino-acid sequence and structural importance of two disulfide bridges. Eur. J. Biochem. 267, 4561–4565. - PubMed
-
- Chen, M.-H., and Citovsky, V. (2003). Systemic movement of a tobamovirus requires host cell pectin methylesterase. Plant J. 35, 386–392. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
