

Assembly of the bacteriophage T4 primosome: single-molecule and ensemble studies
- PMID: 15728347
- PMCID: PMC552937
- DOI: 10.1073/pnas.0500327102
Assembly of the bacteriophage T4 primosome: single-molecule and ensemble studies
Erratum in
- Proc Natl Acad Sci U S A. 2005 Sep 13;102(37):13349-50
Abstract
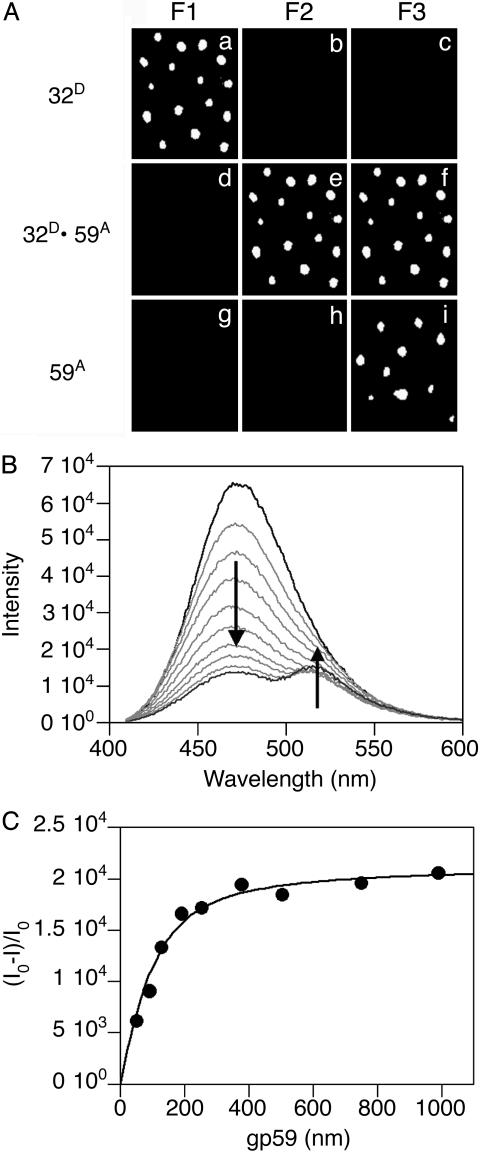
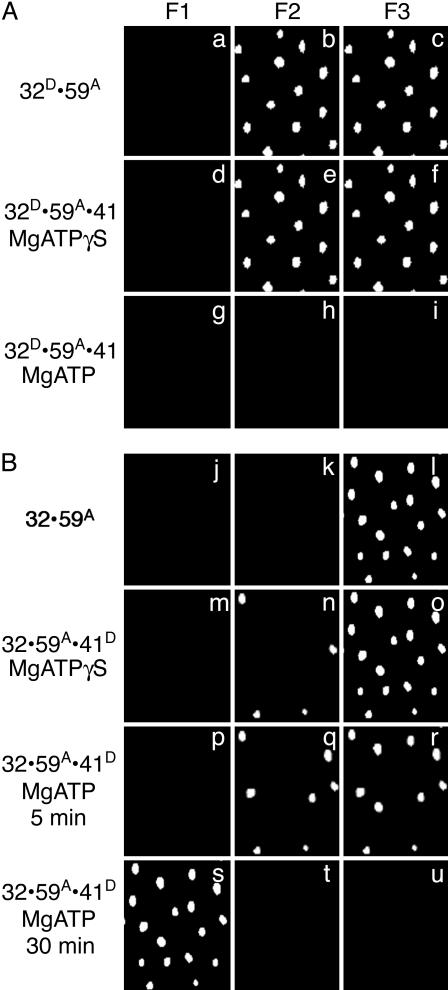
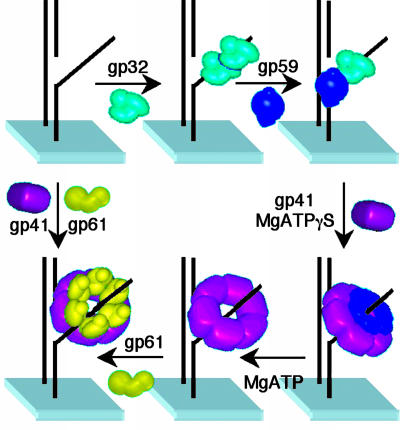
Within replisomes for DNA replication, the primosome is responsible for unwinding double-stranded DNA and synthesizing RNA primers. Assembly of the bacteriophage T4 primosome on individual molecules of ssDNA or forked DNA (fDNA) has been studied by using FRET microscopy. On either DNA substrate, an ordered process of assembly begins with tight 1:1 binding of ssDNA-binding protein (gp32) and helicase-loading protein (gp59) to the DNA. Magnesium adenosine 5'-O-(3-thiotriphosphate) (MgATPgammaS) mediates the weak binding of helicase (gp41) to DNA coated with gp32 and gp59, whereas MgATP induces gp32 and gp59 to dissociate, leaving gp41 bound to the DNA. Finally, primase (gp61) binds to the gp41.DNA complex. Ensemble studies were used to determine protein stoichiometries and binding constants. These single-molecule studies provide an unambiguous description of the pathway for assembly of the primosome on the lagging strand of DNA at a replication fork.
Figures

References
-
- Benkovic, S. J., Valentine, A. M. & Salinas, F. (2001) Annu. Rev. Biochem. 70, 181-208. - PubMed
-
- Yu, X., Jexewska, M. J., Bujalowski, W. & Egelman, E. H. (1996) J. Mol. Biol. 259, 7-14. - PubMed
-
- Keck, J. L. & Berger, J. M. (2001) Nat. Struct. Biol. 8, 2-4. - PubMed
-
- Baker, T. A. & Bell, S. P. (1998) Cell 92, 295-305. - PubMed
-
- Liu, C. C. & Alberts, B. M. (1981) J. Biol. Chem. 256, 2813-2820. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
