

IpaD of Shigella flexneri is independently required for regulation of Ipa protein secretion and efficient insertion of IpaB and IpaC into host membranes
- PMID: 15731041
- PMCID: PMC1064949
- DOI: 10.1128/IAI.73.3.1432-1440.2005
IpaD of Shigella flexneri is independently required for regulation of Ipa protein secretion and efficient insertion of IpaB and IpaC into host membranes
Abstract
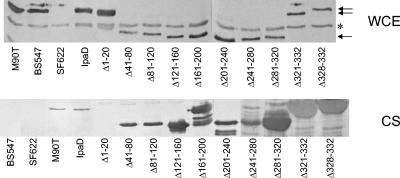
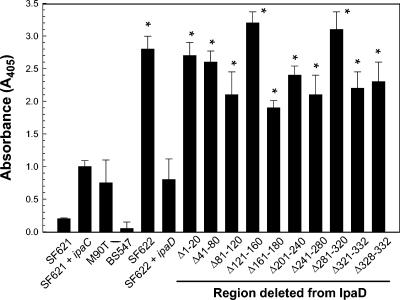
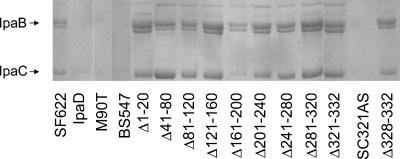
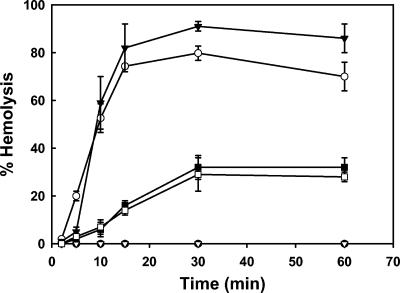
Shigella flexneri causes human dysentery after invading the cells of the colonic epithelium. The best-studied effectors of Shigella entry into colonocytes are the invasion plasmid antigens IpaC and IpaB. These proteins are exported via a type III secretion system (TTSS) to form a pore in the host membrane that may allow the translocation of other effectors into the host cytoplasm. TTSS-mediated secretion of IpaD is also required for translocation pore formation, bacterial invasion, and virulence, but the mechanistic role of this protein is unclear. IpaD is also known to be involved in controlling Ipa protein secretion, but here it is shown that this activity can be separated from its requirement for cellular invasion. Amino acids 40 to 120 of IpaD are not essential for IpaD-dependent invasion; however, deletions in this region still lead to constitutive IpaB/IpaC secretion. Meanwhile, a central deletion causes only a partial loss of control of Ipa secretion but completely eliminates IpaD's invasion function, indicating that IpaD's role in invasion is not a direct outcome of its ability to control Ipa secretion. As shigellae expressing ipaD N-terminal deletion mutations have reduced contact-mediated hemolysis activity and are less efficient at introducing IpaB and IpaC into erythrocyte membranes, it is possible that IpaD is responsible for insertion of IpaB/IpaC pores into target cell membranes. While efficient insertion of IpaB/IpaC pores is needed for optimal invasion efficiency, it may be especially important for Ipa-dependent membrane disruption and thus for efficient vacuolar escape and intercellular spread.
Figures
Similar articles
-
IpaD localizes to the tip of the type III secretion system needle of Shigella flexneri.Infect Immun. 2006 Aug;74(8):4391-400. doi: 10.1128/IAI.00440-06. Infect Immun. 2006. PMID: 16861624 Free PMC article.
-
Identification of functional regions within invasion plasmid antigen C (IpaC) of Shigella flexneri.Mol Microbiol. 2001 Jan;39(1):100-11. doi: 10.1046/j.1365-2958.2001.02210.x. Mol Microbiol. 2001. PMID: 11123692
-
IpaC from Shigella and SipC from Salmonella possess similar biochemical properties but are functionally distinct.Mol Microbiol. 2001 Oct;42(2):469-81. doi: 10.1046/j.1365-2958.2001.02654.x. Mol Microbiol. 2001. PMID: 11703668
-
The roles of the virulence factor IpaB in Shigella spp. in the escape from immune cells and invasion of epithelial cells.Microbiol Res. 2015 Dec;181:43-51. doi: 10.1016/j.micres.2015.08.006. Epub 2015 Sep 2. Microbiol Res. 2015. PMID: 26640051 Review.
-
Molecular and Cellular Mechanisms of Shigella flexneri Dissemination.Front Cell Infect Microbiol. 2016 Mar 11;6:29. doi: 10.3389/fcimb.2016.00029. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27014639 Free PMC article. Review.
Cited by
-
An antibody targeting type III secretion system induces broad protection against Salmonella and Shigella infections.PLoS Negl Trop Dis. 2021 Mar 12;15(3):e0009231. doi: 10.1371/journal.pntd.0009231. eCollection 2021 Mar. PLoS Negl Trop Dis. 2021. PMID: 33711056 Free PMC article.
-
Conformational stability and differential structural analysis of LcrV, PcrV, BipD, and SipD from type III secretion systems.Protein Sci. 2007 Apr;16(4):704-14. doi: 10.1110/ps.062645007. Epub 2007 Feb 27. Protein Sci. 2007. PMID: 17327391 Free PMC article.
-
In vivo versus in vitro protein abundance analysis of Shigella dysenteriae type 1 reveals changes in the expression of proteins involved in virulence, stress and energy metabolism.BMC Microbiol. 2011 Jun 24;11:147. doi: 10.1186/1471-2180-11-147. BMC Microbiol. 2011. PMID: 21702961 Free PMC article.
-
The Type III Secretion Translocation Pore Senses Host Cell Contact.PLoS Pathog. 2016 Mar 29;12(3):e1005530. doi: 10.1371/journal.ppat.1005530. eCollection 2016 Mar. PLoS Pathog. 2016. PMID: 27022930 Free PMC article.
-
Mapping the functional B-cell epitopes of Shigella invasion plasmid antigen D (IpaD).Appl Environ Microbiol. 2024 Aug 21;90(8):e0098824. doi: 10.1128/aem.00988-24. Epub 2024 Jul 31. Appl Environ Microbiol. 2024. PMID: 39082807 Free PMC article.
References
-
- Allaoui, A., P. J. Sansonetti, and C. Parsot. 1993. MxiD, an outer membrane protein necessary for the secretion of the Shigella flexneri Ipa invasins. Mol. Microbiol. 7:59-68. - PubMed
-
- Bennett, J. C. Q., and C. Hughes. 2000. From flagellum assembly to virulence: the extended family of type III export chaperones. Trends Microbiol. 8:202-204. - PubMed
-
- Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 7:248-254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
