

doi: 10.1128/IAI.73.3.1886-1889.2005.
Glutathione and nitrosoglutathione in macrophage defense against Mycobacterium tuberculosis
Affiliations
- PMID: 15731094
- PMCID: PMC1064956
- DOI: 10.1128/IAI.73.3.1886-1889.2005
Item in Clipboard
Glutathione and nitrosoglutathione in macrophage defense against Mycobacterium tuberculosis
Infect Immun.
2005 Mar.
Abstract
We demonstrate that Mycobacterium tuberculosis grown in vitro is sensitive to glutathione and its derivative S-nitrosoglutathione. Furthermore, our infection studies with J774.1 macrophages indicate that glutathione is essential for the control of the intracellular growth of M. tuberculosis. This study indicates the important role of glutathione in the control of macrophages by M. tuberculosis.
Figures

Growth of M. tuberculosis in the presence and absence of GSH (A) and GSNO (B) as determined by measuring OD. The H37Rv (Rv) cell suspension was treated with 5 mM GSH or 5 mM GSNO or left untreated. The OD was measured every day for 5 days (d.) (A). All experiments were repeated three times in triplicate.

Growth of H37Rv in untreated (A) and IFN-γ-LPS-treated (B) J774.1 cells. Experiments with H37Rv-infected macrophages, maintained in the absence (A) and presence (B) of IFN-γ plus LPS, were terminated at 1 and 72 h to determine the growth of H37Rv inside J774.1 cells. * denotes a statistically significant increase in the number of CFU between 1 and 72 h (P < 0.0006). Data are averages of results from six different experiments performed in triplicate.
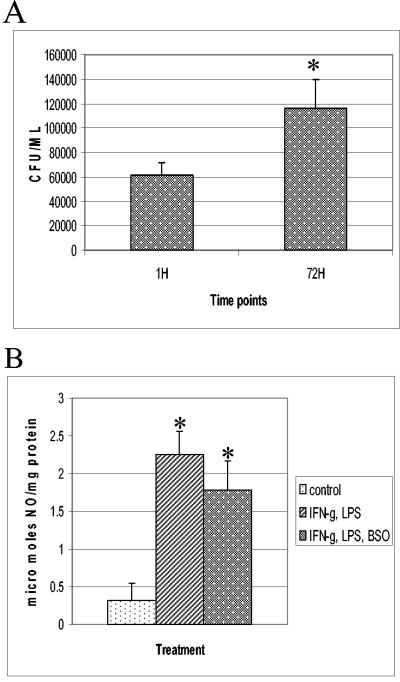
(A) Growth of H37Rv in IFN-γ-LPS-BSO-treated J774.1 cells. Experiments with H37Rv-infected macrophages treated with IFN-γ plus LPS and BSO were terminated at 1 and 72 h to determine the levels of growth of H37Rv inside J774.1 cells. Statistical significance was calculated with the Statview program. * represents a statistically significant increase in the number of CFU between 1 and 72 h (P < 0.0083). Data are averages of results from from six different experiments performed in triplicate. (B) NO estimation in J774.1 cells. Nitrite levels in macrophage supernatants were determined spectrophotometrically by a Greiss reaction. Data are averages of results from five different experiments. * indicates a statistically significant increase in nitrite levels between control and IFN-γ-LPS- or IFN-γ-LPS-BSO-treated macrophages. For values for the control versus those after IFN-γ-LPS treatment, P was <0.0001. For values for the control versus those after IFN-γ-LPS-BSO treatment, P was <0.0030.

Estimation of GSH levels in J774.1 cells by flow cytometry (A) and spectrophotometry (B). (A) GSH was quantitated in J774.1 cells by staining with monochlorobimane. * signifies a statistically significant decrease in the number of fluorescent cells in IFN-γ-LPS-BSO-treated macrophages compared to that for control or IFN-γ-LPS-treated macrophages. For values for the control versus those after IFN-γ-LPS-BSO treatment, P was <0.0001. For values after IFN-γ-LPS treatment versus those after IFN-γ-LPS-BSO treatment, P was <0.0026. Data are averages of results from three different experiments. (B) GSH was also assayed by spectrophotometry. Data are averages from three different experiments. * designates a statistically significant decrease in intracellular GSH levels in IFN-γ-LPS-BSO-treated macrophages compared to those in control macrophages or IFN-γ-LPS-treated macrophages. For values for the control versus those after IFN-γ-LPS—BSO treatment, P was <0.0024. For values after IFN-γ-LPS treatment versus IFN-γ-LPS-BSO treatment, P was <0.0028.
Similar articles
-
Characterization of a glutathione metabolic mutant of Mycobacterium tuberculosis and its resistance to glutathione and nitrosoglutathione.J Bacteriol. 2006 Feb;188(4):1364-72. doi: 10.1128/JB.188.4.1364-1372.2006. J Bacteriol. 2006. PMID: 16452418 Free PMC article.
-
Macrophage infection models for Mycobacterium tuberculosis.Methods Mol Biol. 2015;1285:329-41. doi: 10.1007/978-1-4939-2450-9_20. Methods Mol Biol. 2015. PMID: 25779326
-
Development of infection model for studying intracellular gene expression of Mycobacterium tuberculosis.Afr J Med Med Sci. 2009 Dec;38(4):325-32. Afr J Med Med Sci. 2009. PMID: 20499625
-
Killing intracellular mycobacteria in in vitro macrophage systems: what may be the role of known host microbicidal mechanisms?Res Microbiol. 1990 Feb;141(2):217-30. doi: 10.1016/0923-2508(90)90034-n. Res Microbiol. 1990. PMID: 2111923 Review. No abstract available.
-
[Role of immune responses in tuberculosis].Kekkaku. 1997 Dec;72(12):689-92. Kekkaku. 1997. PMID: 9556383 Review. Japanese. No abstract available.
Cited by
-
The Effects of Oral Liposomal Glutathione and In Vitro Everolimus in Altering the Immune Responses against Mycobacterium bovis BCG Strain in Individuals with Type 2 Diabetes.Biomol Concepts. 2021 May 9;12(1):16-26. doi: 10.1515/bmc-2021-0003. Biomol Concepts. 2021. PMID: 33966361 Free PMC article. Clinical Trial.
-
Host-directed therapy for tuberculosis.Eur J Med Res. 2025 Apr 11;30(1):267. doi: 10.1186/s40001-025-02443-4. Eur J Med Res. 2025. PMID: 40211397 Free PMC article. Review.
-
Additive Effects of Glutathione in Improving Antibiotic Efficacy in HIV-M.tb Co-Infection in the Central Nervous System: A Systematic Review.Viruses. 2025 Jan 17;17(1):127. doi: 10.3390/v17010127. Viruses. 2025. PMID: 39861915 Free PMC article.
-
Investigating the causes for decreased levels of glutathione in individuals with type II diabetes.PLoS One. 2015 Mar 19;10(3):e0118436. doi: 10.1371/journal.pone.0118436. eCollection 2015. PLoS One. 2015. PMID: 25790445 Free PMC article.
-
Liposomal Glutathione Helps to Mitigate Mycobacterium tuberculosis Infection in the Lungs.Antioxidants (Basel). 2022 Mar 30;11(4):673. doi: 10.3390/antiox11040673. Antioxidants (Basel). 2022. PMID: 35453358 Free PMC article.
References
-
- Adams, L. B., M. C. Dinauer, D. E. Morgenstern, and J. L. Krahenbuhl. 1997. Comparison of the roles of reactive oxygen and nitrogen intermediates in the host response to Mycobacterium tuberculosis using transgenic mice. Tuber. Lung Dis. 78:237-246. - PubMed
-
- Anderson, M. T., M. Roederer, I. Tjioe, and L. A. Herzenberg. 1996. p. 1-9. In L. A. Herzenberg, C. Blackwell, and D. Weir (ed.), The handbook of experimental immunology, 5th ed., vol. 1B. Blackwell Scientific, Boston, Mass.
-
- Bloom, B. R., and C. J. Murray. 1992. Tuberculosis: commentary on a reemergent killer. Science 257:1055-1064. - PubMed
-
- Chan, E. D., J. Chan, and N. W. Schluger. 2001. What is the role of nitric oxide in murine and human host defense against tuberculosis? Current knowledge. Am. J. Respir. Cell. Mol. Biol. 25:606-612. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
