

Deletion of highly conserved arginine-rich RNA binding motif in cowpea chlorotic mottle virus capsid protein results in virion structural alterations and RNA packaging constraints
- PMID: 15731222
- PMCID: PMC1075711
- DOI: 10.1128/JVI.79.6.3277-3288.2005
Deletion of highly conserved arginine-rich RNA binding motif in cowpea chlorotic mottle virus capsid protein results in virion structural alterations and RNA packaging constraints
Abstract
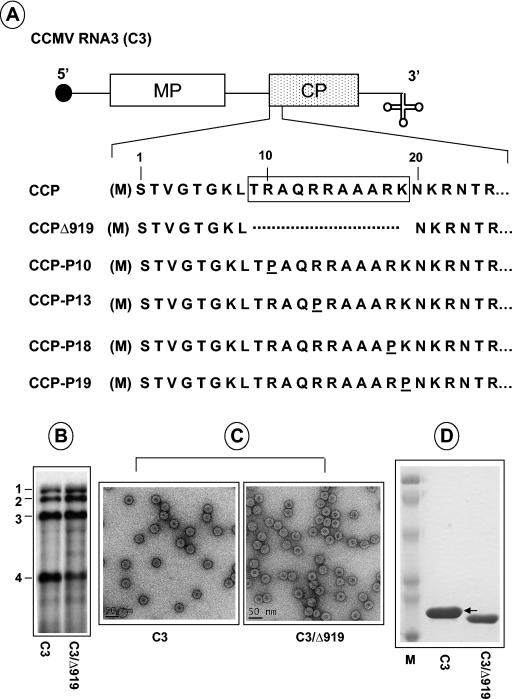
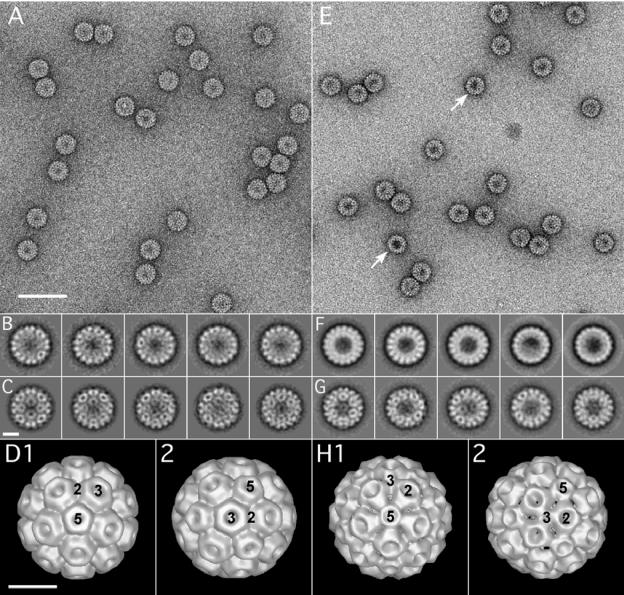
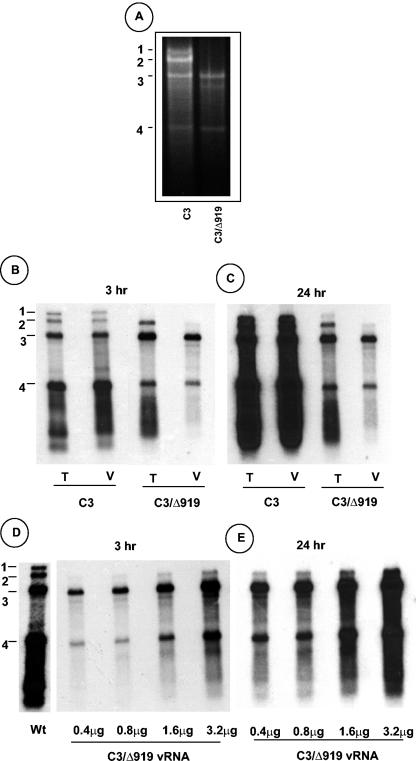
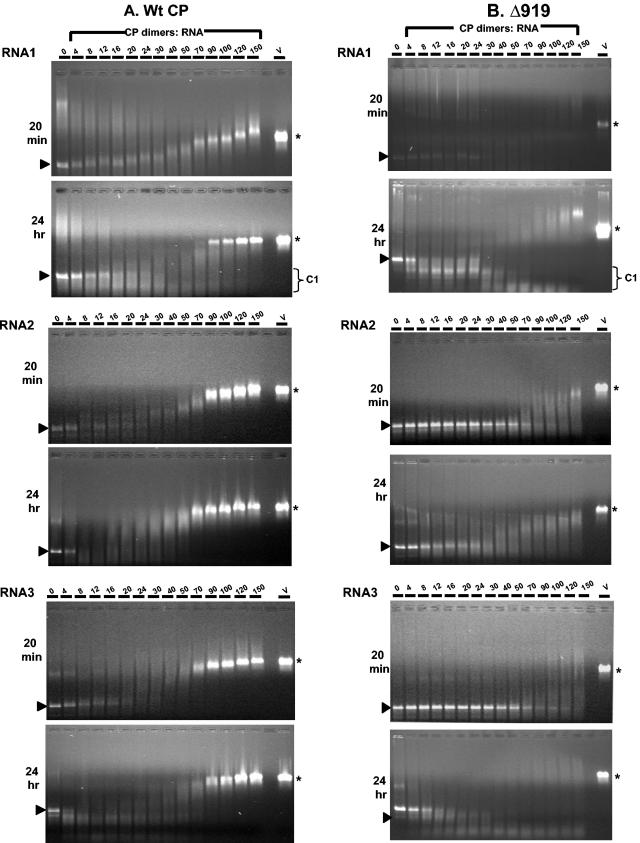
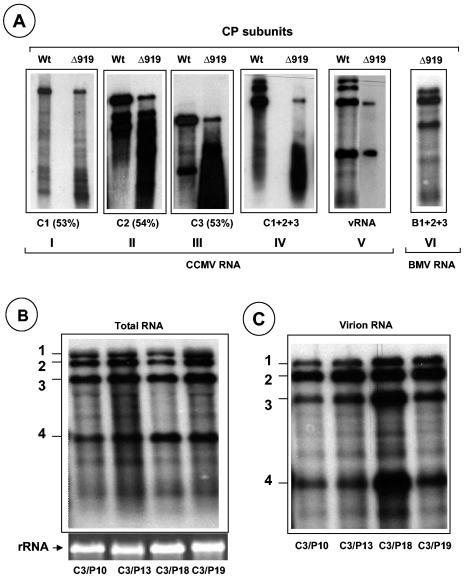
The N-proximal region of cowpea chlorotic mottle virus (CCMV) capsid protein (CP) contains an arginine-rich RNA binding motif (ARM) that is also found in the CPs of other members of Bromoviridae and in other RNA binding proteins such as the Tat and Rev proteins of human immunodeficiency virus. To assess the critical role played by this motif during encapsidation, a variant of CCMV RNA3 (C3) precisely lacking the ARM region (C3/Delta919) of its CP gene was constructed. The biology and the competence of the matured CP derived in vivo from C3/Delta919 to assemble and package progeny RNA was examined in whole plants. Image analysis and computer-assisted three-dimensional reconstruction of wild-type and mutant virions revealed that the CP subunits bearing the engineered deletion assembled into polymorphic virions with altered surface topology. Northern blot analysis of virion RNA from mutant progeny demonstrated that the engineered mutation down-regulated packaging of all four viral RNAs; however, the packaging effect was more pronounced on genomic RNA1 and RNA2 than genomic RNA3 and its CP mRNA. In vitro assembly assays with mutant CP subunits and RNA transcripts demonstrated that the mutant CP is inherently not defective in packaging genomic RNA1 (53%) and RNA2 (54%), but their incorporation into virions was competitively inhibited by the presence of other viral RNAs. Northern blot analysis of RNA encapsidation in vivo of two distinct bromovirus RNA3 chimeras, constructed by exchanging CPs having the Delta919 deletion, demonstrated that the role of the conserved N-terminal ARM in recognizing and packaging specific RNA is distinct for each virus.
Figures
References
-
- Bancroft, J. B. 1970. The self-assembly of spherical plant viruses. Adv. Virus Res. 16:99-134. - PubMed
-
- Böttcher, B., S. A. Wynne, and R. A. Crowther. 1997. Determination of the fold of the core protein of hepatitis B virus by electron cryomicroscopy. Nature 386:88-91. - PubMed
-
- Burd, C. G., and G. Dreyfuss. 1994. Conserved structures and diversity of functions of RNA-binding proteins. Science 265:615-621. - PubMed
-
- Choi, Y. G., and A. L. N. Rao. 2000. Molecular studies on bromovirus capsid protein. VII. Selective packaging of BMV RNA4 by specific N-terminal arginine residues. Virology 275:207-217. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
