

Differential role for TLR3 in respiratory syncytial virus-induced chemokine expression
- PMID: 15731229
- PMCID: PMC1075725
- DOI: 10.1128/JVI.79.6.3350-3357.2005
Differential role for TLR3 in respiratory syncytial virus-induced chemokine expression
Abstract
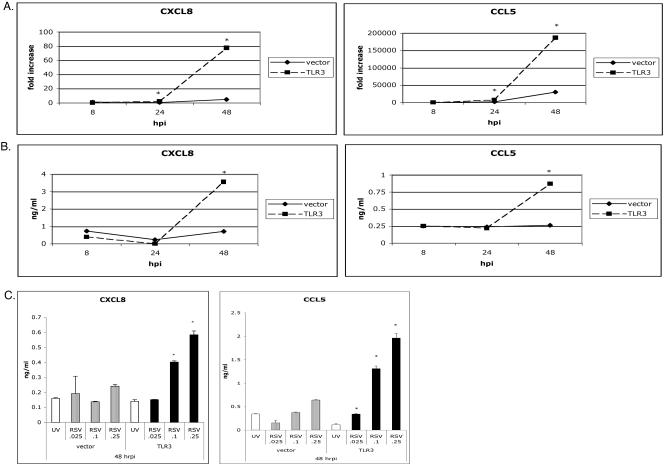
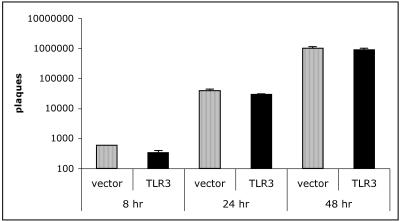
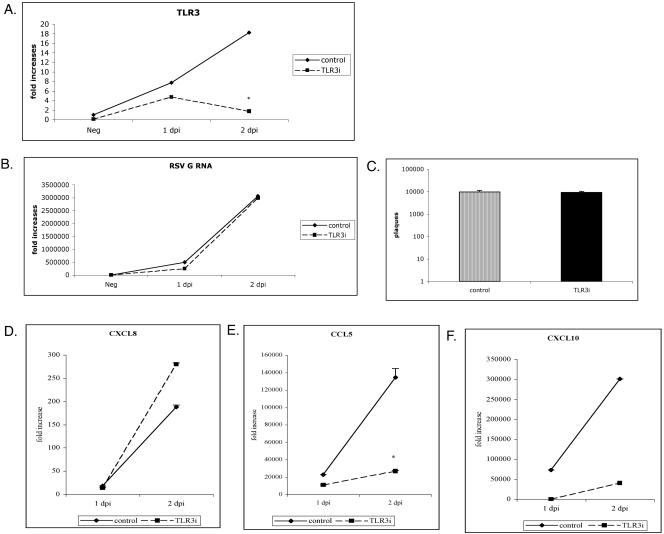
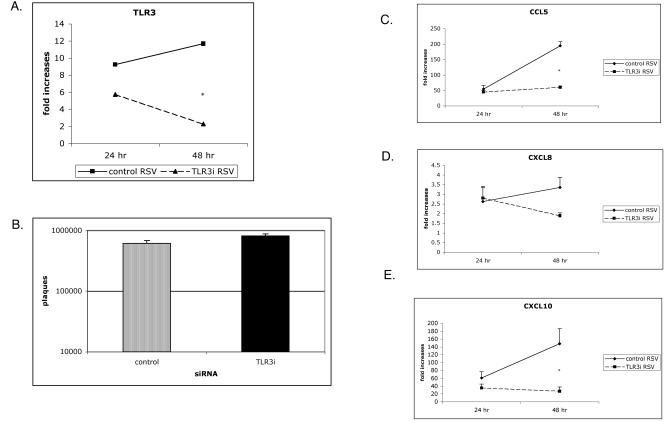
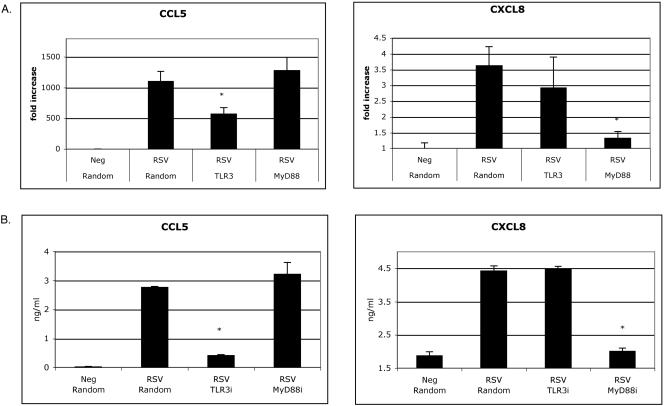
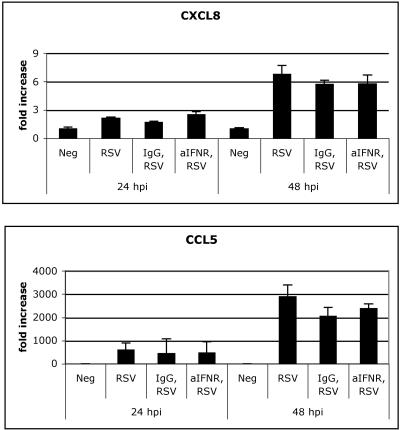
Respiratory syncytial virus (RSV) is the leading cause of lower respiratory tract infection in young infants worldwide. Previous studies have reported that the induction of interleukin-8/CXCL8 and RANTES/CCL5 correlates with disease severity in humans. The production of these chemokines is elicited by viral replication and is NF-kappaB dependent. RSV, a negative-sense single-stranded RNA virus, requires full-length positive-sense RNA for synthesis of new viral RNA. The aim of our studies was to investigate whether active viral replication by RSV could evoke chemokine production through TLR3-mediated signaling pathways. In TLR3-transfected HEK 293 cells, live RSV preferentially activated chemokines in both a time- and dose-dependent manner compared to vector controls. RSV was also shown to upregulate TLR3 in human lung fibroblasts and epithelial cells (MRC-5 and A549). Targeting the expression of TLR3 with small interfering RNA decreased synthesis of IP-10/CXCL10 and CCL5 but did not significantly reduce levels of CXCL8. Blocking the expression of the adapter protein MyD88 established a role for MyD88 in CXCL8 production, whereas CCL5 synthesis was found to be MyD88 independent. Production of CCL5 by RSV was induced directly through TLR3 signaling pathways and did not require interferon (IFN) signaling through the IFN-alpha/beta receptor. TLR3 did not affect viral replication, since equivalent viral loads were recovered from RSV-infected cells despite altered TLR3 expression. Taken together, our studies indicate that TLR3 mediates inflammatory cytokine and chemokine production in RSV-infected epithelial cells.
Figures
Similar articles
-
Synergistic induction of CXCL9 and CXCL11 by Toll-like receptor ligands and interferon-gamma in fibroblasts correlates with elevated levels of CXCR3 ligands in septic arthritis synovial fluids.J Leukoc Biol. 2004 May;75(5):777-84. doi: 10.1189/jlb.1003524. Epub 2004 Mar 2. J Leukoc Biol. 2004. PMID: 14996826
-
Inhibition of respiratory syncytial virus infection with the CC chemokine RANTES (CCL5).J Med Virol. 2004 Jun;73(2):300-8. doi: 10.1002/jmv.20091. J Med Virol. 2004. PMID: 15122808
-
Respiratory syncytial virus-induced chemokine production: linking viral replication to chemokine production in vitro and in vivo.J Infect Dis. 2004 Apr 15;189(8):1419-30. doi: 10.1086/382958. Epub 2004 Apr 2. J Infect Dis. 2004. PMID: 15073679
-
Toll-like receptor 3: a link between toll-like receptor, interferon and viruses.Microbiol Immunol. 2004;48(3):147-54. doi: 10.1111/j.1348-0421.2004.tb03500.x. Microbiol Immunol. 2004. PMID: 15031527 Review.
-
Immunity Cell Responses to RSV and the Role of Antiviral Inhibitors: A Systematic Review.Infect Drug Resist. 2022 Dec 14;15:7413-7430. doi: 10.2147/IDR.S387479. eCollection 2022. Infect Drug Resist. 2022. PMID: 36540102 Free PMC article. Review.
Cited by
-
Genetic replacement of surfactant protein-C reduces respiratory syncytial virus induced lung injury.Respir Res. 2013 Feb 12;14(1):19. doi: 10.1186/1465-9921-14-19. Respir Res. 2013. PMID: 23399055 Free PMC article.
-
Basic research on virus-induced asthma exacerbation: inhibition of inflammatory chemokine expression by fluticasone propionate.Int Arch Allergy Immunol. 2013;161 Suppl 2(Suppl 2):84-92. doi: 10.1159/000350455. Epub 2013 May 29. Int Arch Allergy Immunol. 2013. PMID: 23711858 Free PMC article.
-
Assessing the protection elicited by virus-like particles expressing the RSV pre-fusion F and tandem repeated G proteins against RSV rA2 line19F infection in mice.Respir Res. 2024 Jan 4;25(1):7. doi: 10.1186/s12931-023-02641-w. Respir Res. 2024. PMID: 38178222 Free PMC article.
-
Inhibition of respiratory syncytial virus replication and suppression of RSV-induced airway inflammation in neonatal rats by colchicine.3 Biotech. 2019 Nov;9(11):392. doi: 10.1007/s13205-019-1917-z. Epub 2019 Oct 10. 3 Biotech. 2019. PMID: 31656730 Free PMC article.
-
Cigarette smoke dampens antiviral signaling in small airway epithelial cells by disrupting TLR3 cleavage.Am J Physiol Lung Cell Mol Physiol. 2018 Mar 1;314(3):L505-L513. doi: 10.1152/ajplung.00406.2017. Epub 2017 Dec 14. Am J Physiol Lung Cell Mol Physiol. 2018. PMID: 29351447 Free PMC article.
References
-
- Alexopoulou, L., A. C. Holt, R. Medzhitov, and R. A. Flavell. 2001. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 413:732-738. - PubMed
-
- Atreya, P. L., and S. Kulkarni. 1999. Respiratory syncytial virus strain A2 is resistant to the antiviral effects of type I interferons and human MxA. Virology 261:227-241. - PubMed
-
- Becker, M. N., G. Diamond, M. W. Verghese, and S. H. Randell. 2000. CD14-dependent lipopolysaccharide-induced beta-defensin-2 expression in human tracheobronchial epithelium. J. Biol. Chem. 275:29731-29736. - PubMed
-
- Casola, A., N. Burger, T. Liu, M. Jamaluddin, A. R. Brasier, and R. P. Garofalo. 2001. Oxidant tone regulates RANTES gene expression in airway epithelial cells infected with respiratory syncytial virus. Role in viral-induced interferon regulatory factor activation. J. Biol. Chem. 276:19715-19722. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
