

Importance of both the coding and the segment-specific noncoding regions of the influenza A virus NS segment for its efficient incorporation into virions
- PMID: 15731270
- PMCID: PMC1075679
- DOI: 10.1128/JVI.79.6.3766-3774.2005
Importance of both the coding and the segment-specific noncoding regions of the influenza A virus NS segment for its efficient incorporation into virions
Abstract
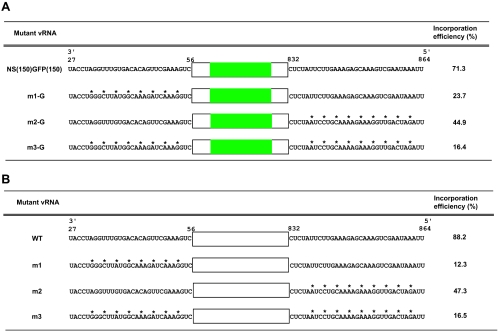
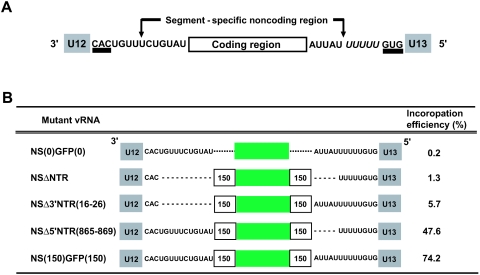
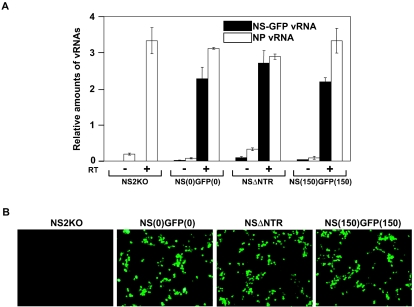
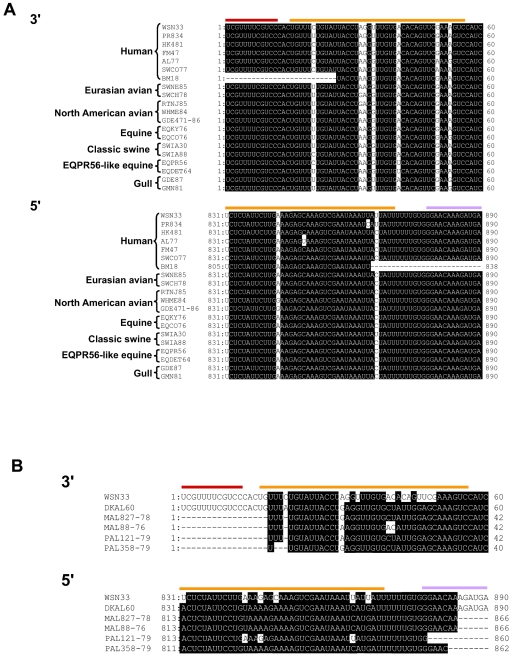
The genome of influenza A virus consists of eight single-strand negative-sense RNA segments, each comprised of a coding region and a noncoding region. The noncoding region of the NS segment is thought to provide the signal for packaging; however, we recently showed that the coding regions located at both ends of the hemagglutinin and neuraminidase segments were important for their incorporation into virions. In an effort to improve our understanding of the mechanism of influenza virus genome packaging, we sought to identify the regions of NS viral RNA (vRNA) that are required for its efficient incorporation into virions. Deletion analysis showed that the first 30 nucleotides of the 3' coding region are critical for efficient NS vRNA incorporation and that deletion of the 3' segment-specific noncoding region drastically reduces NS vRNA incorporation into virions. Furthermore, silent mutations in the first 30 nucleotides of the 3' NS coding region reduced the incorporation efficiency of the NS segment and affected virus replication. These results suggested that segment-specific noncoding regions together with adjacent coding regions (especially at the 3' end) form a structure that is required for efficient influenza A virus vRNA packaging.
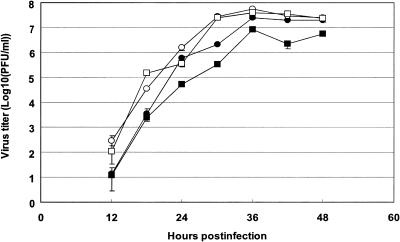
Figures
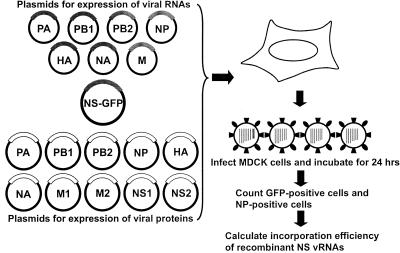
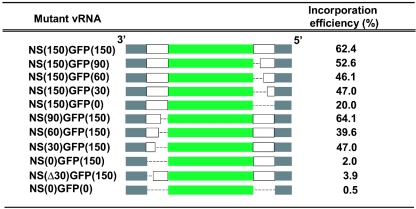
References
-
- Baez, M., J. J. Zazra, R. M. Elliott, J. F. Young, and P. Palese. 1981. Nucleotide sequence of the influenza A/duck/Alberta/60/76 virus NS RNA: conservation of the NS1/NS2 overlapping gene structure in a divergent influenza virus RNA segment. Virology 113:397-402. - PubMed
-
- Chen, W., P. A. Calvo, D. Malide, J. Gibbs, U. Schubert, I. Bacik, S. Basta, R. O'Neill, J. Schickli, P. Palese, P. Henklein, J. R. Bennink, and J. W. Yewdell. 2001. A novel influenza A virus mitochondrial protein that induces cell death. Nat. Med. 7:1306-1312. - PubMed
-
- Enami, M., R. Fukuda, and A. Ishihama. 1985. Transcription and replication of eight RNA segments of influenza virus. Virology 142:68-77. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
