

Endocytosis of the Nipah virus glycoproteins
- PMID: 15731282
- PMCID: PMC1075720
- DOI: 10.1128/JVI.79.6.3865-3872.2005
Endocytosis of the Nipah virus glycoproteins
Abstract
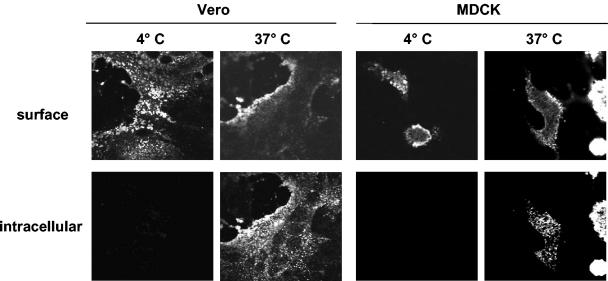
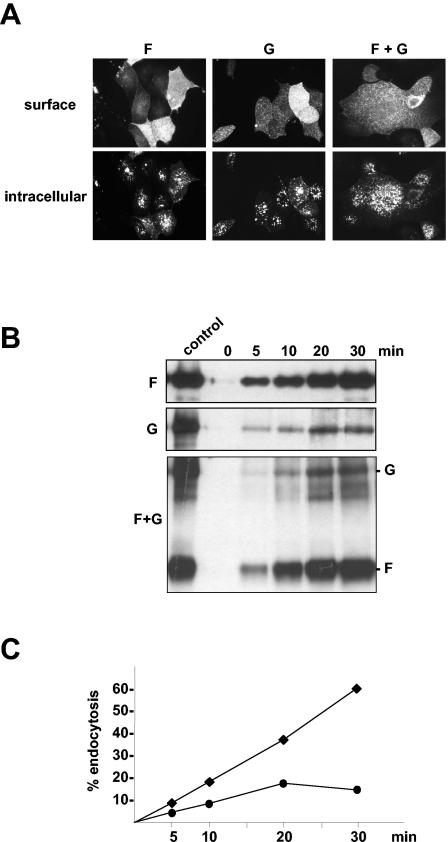
Nipah virus (NiV), a highly pathogenic member of the family Paramyxoviridae, encodes the surface glycoproteins F and G. Since internalization of the NiV envelope proteins from the cell surface might be of functional importance for viral pathogenesis either by regulating cytopathogenicity or by modulating recognition of infected cells by the immune system, we analyzed the endocytosis of the NiV F and G proteins. Interestingly, we found both glycoproteins to be internalized in infected and transfected cells. As endocytosis is normally mediated by tyrosine- or dileucine-dependent signals in the cytoplasmic tails of transmembrane proteins, all potential internalization signals in the NiV glycoproteins were mutated. Whereas the G protein appeared to be constitutively internalized with the bulk flow during membrane turnover, uptake of the F protein was found to be signal mediated. F endocytosis clearly depended on a membrane-proximal YXXPhi motif and was found to be of functional importance for the biological activity of the protein.
Figures

References
-
- Berlioz-Torrent, C., B. L. Shacklett, L. Erdtmann, L. Delamarre, I. Bouchaert, P. Sonigo, M. C. Dokhelar, and R. Benarous. 1999. Interactions of the cytoplasmic domains of human and simian retroviral transmembrane proteins with components of the clathrin adaptor complexes modulate intracellular and cell surface expression of envelope glycoproteins. J. Virol. 73:1350-1361. - PMC - PubMed
-
- Bonifacino, J. S., and L. M. Traub. 2003. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu. Rev. Biochem. 72:395-447. - PubMed
-
- Chua, K. B., K. J. Goh, K. T. Wong, A. Kamarulzaman, P. S. Tan, T. G. Ksiazek, S. R. Zaki, G. Paul, S. K. Lam, and C. T. Tan. 1999. Fatal encephalitis due to Nipah virus among pig-farmers in Malaysia. Lancet 354:1257-1259. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
