

Double insertions of extraocular rectus muscles in humans and the pulley theory
- PMID: 15733302
- PMCID: PMC1571479
- DOI: 10.1111/j.1469-7580.2005.00383.x
Double insertions of extraocular rectus muscles in humans and the pulley theory
Abstract
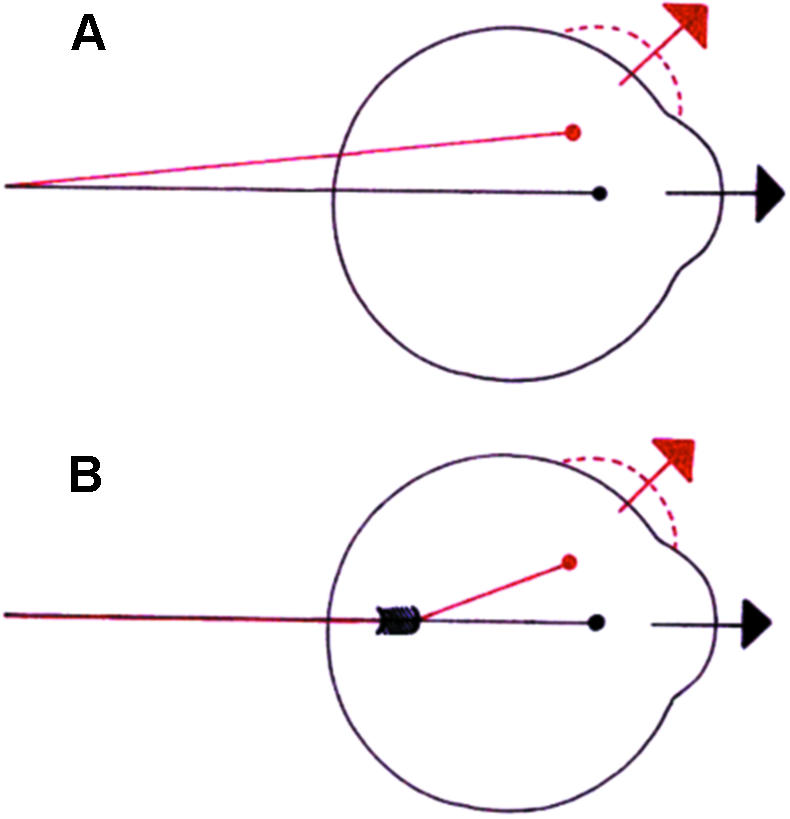
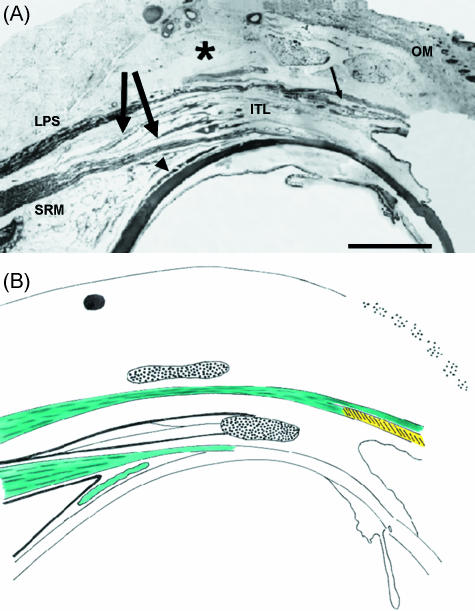
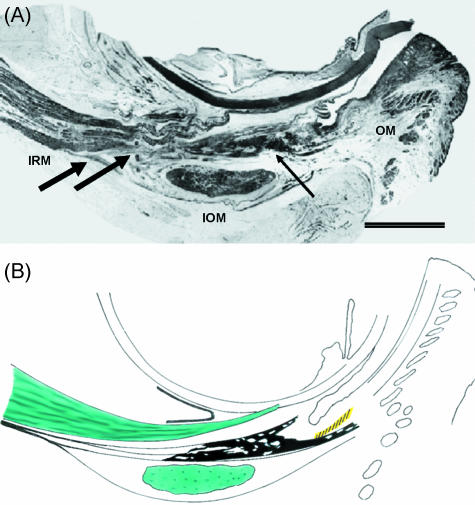
Recent studies have promoted the concept that rectus muscles pass through connective tissue pulleys located near the equator of the eye and act, in effect, as the muscle origins. Orbital muscle fibres (facing bone) terminate in pulleys, permitting adjustment of their position independent of the global fibres responsible for rotating the eye. The structure of pulleys (or muscle sleeves) and the passage taken by their muscle fibre insertions are unclear, and a detailed description is presented here together with a review of the active pulley hypothesis. Segments including the full width of single muscles were removed from the full orbital contents of dissection room cadavers and fresh perfusion-fixed rhesus and cynomolgus monkeys and prepared for light microscopy. Thin longitudinal sections were cut as facets from resin-embedded tissue blocks and montages assembled. Interrupted serial sections of selected regions of both species and ultrathin sections of monkey material were prepared for light and electron microscopy, respectively. Slender tendons leave the orbital surface of rectus muscles at intervals, aggregating and entering sleeves in humans and monkey; less frequently, tendons pass from the global surface to sleeves or insert directly in the posterior fascia bulbi. The orbital sides of sleeve rings are continuous with the fascial canopy of the globe and are 5-6 times as thick as the global sides; sleeve structure differs in the four recti. Medial rectus sleeves are the thickest, and contain smooth muscle, whereas little or none is present in the other rectus sleeves. Superior rectus sleeves are variable in structure and relatively insubstantial. A narrow interval separates muscles from the surrounding connective tissue equatorially in some preparations, consistent with a capacity to slide, but the tissues are contiguous in others, especially in monkey material. The structural organization of sleeves and their tendons, together with other presented factors, is inconsistent with a facility for the separate adjustment of sleeve position. The results favour the theory that sleeve tendons have just one role, to counter the viscoelastic resistance of global fascia - ocular and sleeve muscle fibres acting in unison. Whether the fragile sleeve structure can meet the physical demands of pulleys is questionable; but otherwise the veracity of the pulley hypothesis cannot be assessed from the structural relations of muscles and fascia bulbi reported.
Figures



References
-
- Acheson GH. The topographical anatomy of the smooth muscle of the cat's nictitating membrane. Anat. Rec. 1938;71:297–311.
-
- Adachi B. Das Knorpelstück in der Plica semilunaris conjunctivae der Japaner. Z. Morph. Anthrop. 1906;9:325–326.
-
- Bartels P. Histolisch-anthropologische Untersuchungen der Plica semilunaris bei Herero und Hottentotten, sowie bei einigen Anthropoiden. Arch. Mikrosk. Anat. 1. Abt. 1911;78:29–564.
-
- Clark RA, Miller JM, Demer JL. Three-dimensional location of human rectus pulleys by path inflections in secondary gaze positions. Invest. Ophthalmol. Vis. Sci. 2000;41:3787–3797. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
