

A genome-wide analysis of the effects of sucrose on gene expression in Arabidopsis seedlings under anoxia
- PMID: 15734908
- PMCID: PMC1065412
- DOI: 10.1104/pp.104.057299
A genome-wide analysis of the effects of sucrose on gene expression in Arabidopsis seedlings under anoxia
Abstract
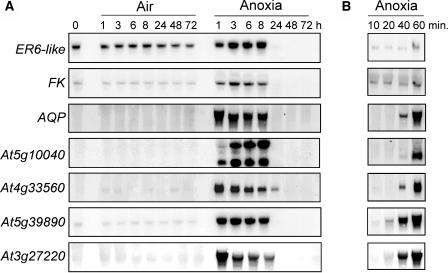
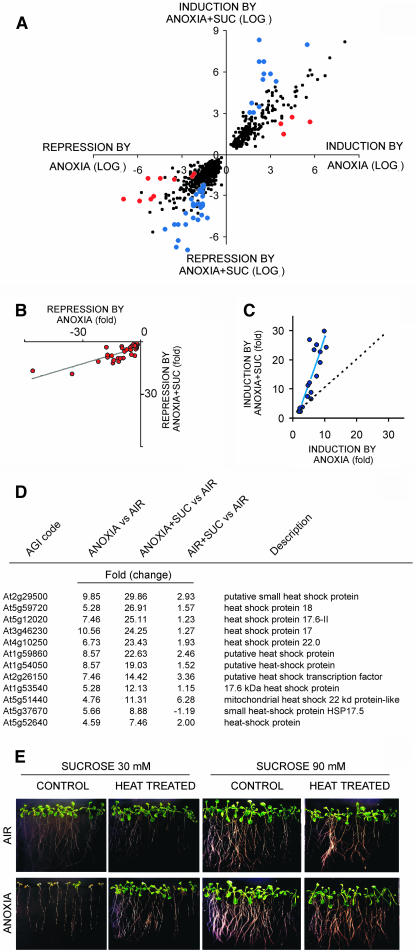
Exogenous sucrose (Suc) greatly enhances anoxia tolerance of Arabidopsis (Arabidopsis thaliana) seedlings. We used the Affymetrix ATH1 GeneChip containing more than 22,500 probe sets to explore the anaerobic transcriptome of Arabidopsis seedlings kept under anoxia for 6 h in presence or absence of exogenous Suc. Functional clustering was performed using the MapMan software. Besides the expected induction of genes encoding enzymes involved in Suc metabolism and alcoholic fermentation, a large number of genes not related to these pathways were affected by anoxia. Addition of exogenous Suc mitigated the effects of anoxia on auxin responsive genes that are repressed under oxygen deprivation. Anoxia-induced Suc synthases showed a lower induction in presence of exogenous Suc, suggesting that induction of these genes might be related to an anoxia-dependent sugar starvation. Anoxic induction of genes coding for heat shock proteins was much stronger in presence of exogenous Suc. Interestingly, a short heat treatment enhanced anoxia tolerance, suggesting that heat shock proteins may play a role in survival to low oxygen. These results provide insight into the effects of Suc on the anoxic transcriptome and provide a list of candidate genes that enhance anoxia tolerance of Suc-treated seedlings.
Figures



References
-
- Baud S, Vaultier MN, Rochat C (2004) Structure and expression profile of the sucrose synthase multigene family in Arabidopsis. J Exp Bot 55: 397–409 - PubMed
-
- Dolferus R, Ellis M, de Bruxelles G, Trevaskis B, Hoeren F, Dennis ES, Peacock WJ (1997) Strategies of gene action in Arabidopsis during hypoxia. Ann Bot (Lond) 79: 21–31
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
