

Disruption of lateral olivocochlear neurons via a dopaminergic neurotoxin depresses sound-evoked auditory nerve activity
- PMID: 15735934
- PMCID: PMC2504639
- DOI: 10.1007/s10162-004-5009-2
Disruption of lateral olivocochlear neurons via a dopaminergic neurotoxin depresses sound-evoked auditory nerve activity
Abstract
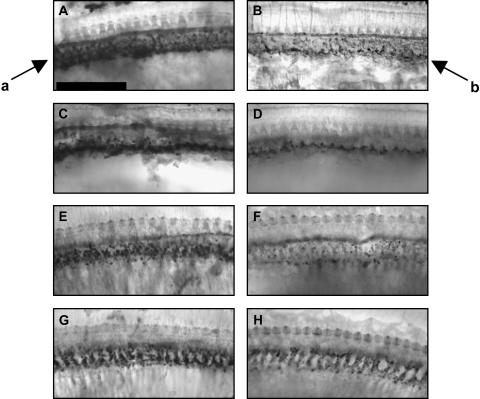
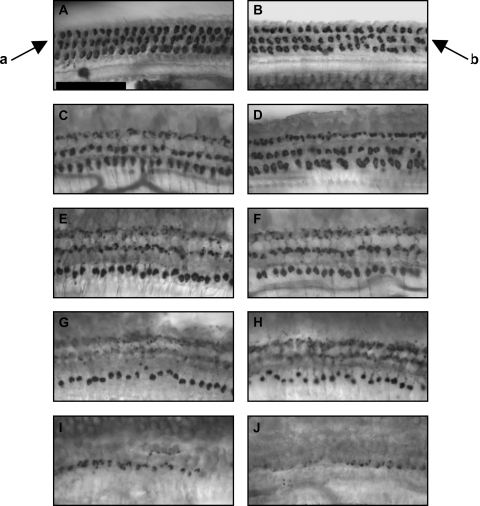
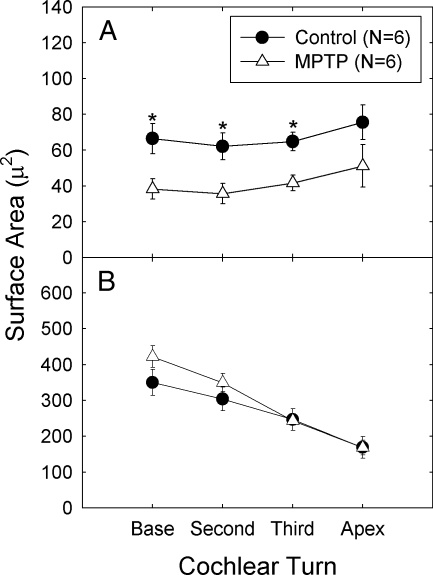
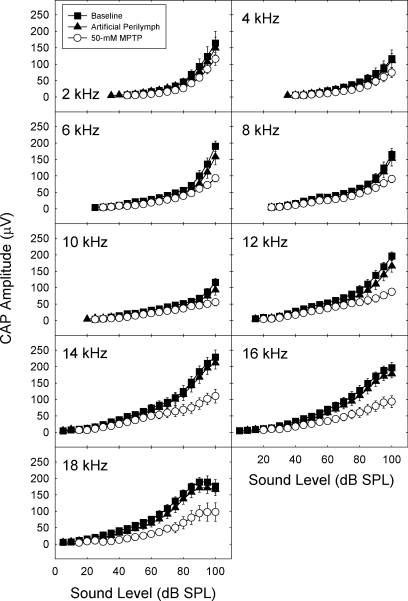
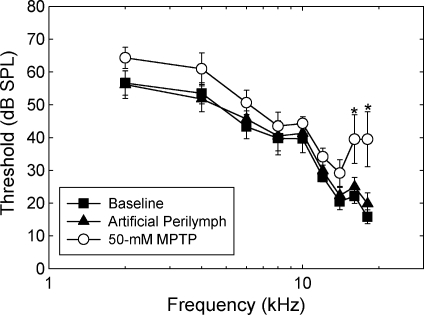
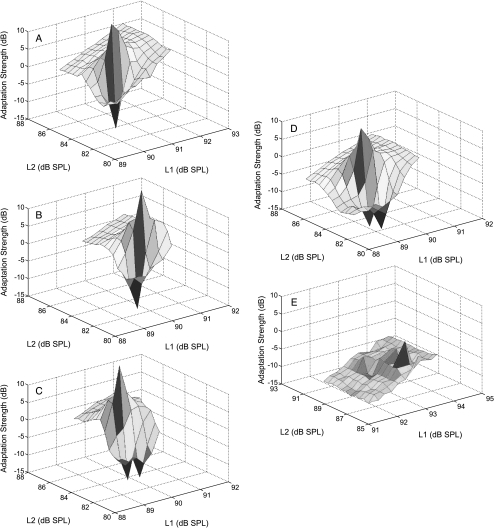
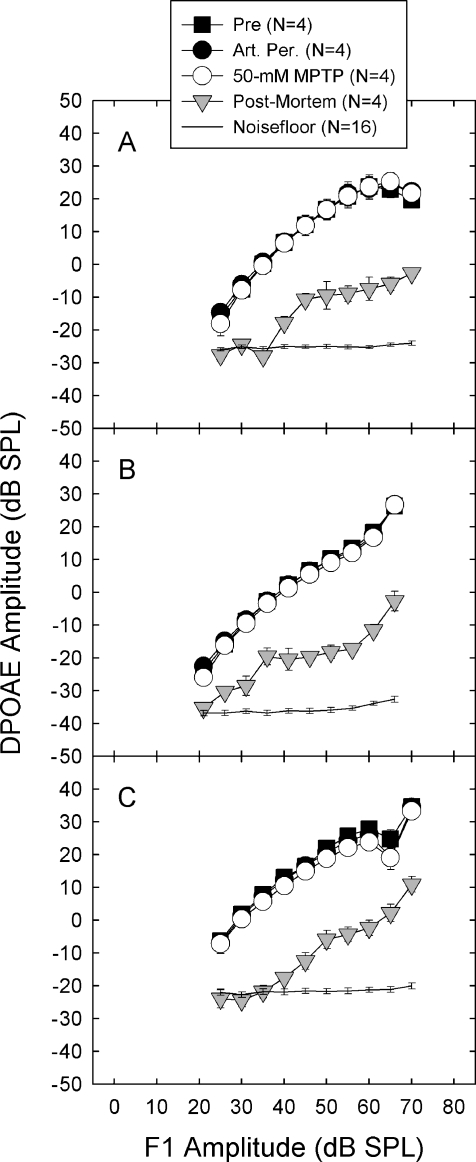
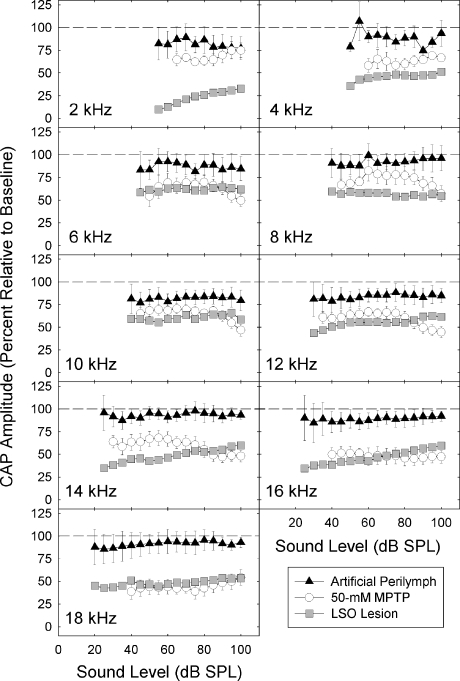
We applied the dopaminergic (DA) neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) to the guinea pig cochlear perilymph. Immunolabeling of lateral olivocochlear (LOC) neurons using antibodies against synaptophysin was reduced after the MPTP treatment. In contrast, labeling of the medial olivocochlear innervation remained intact. As after brainstem lesions of the lateral superior olive (LSO), the site of origin of the LOC neurons, the main effect of disrupting LOC innervation of the cochlea via MPTP was a depression of the amplitude of the compound action potential (CAP). CAP amplitude depression was similar to that produced by LSO lesions. Latency of the N1 component of the CAP, and distortion product otoacoustic emission amplitude and adaptation were unchanged by the MPTP treatment. This technique for selectively lesioning descending LOC efferents provides a new opportunity for examining LOC modulation of afferent activity and behavioral measures of perception.
Figures
References
-
- Abou-Madi L, Pontarotti P, Tramu G, Cupo A, Eybalin M. Coexistence of putative neuroactive substances in lateral olivocochlear neurons of rat and guinea pig. Hear. Res. 1987;30:135–146. - PubMed
-
- Adams JC, Mroz EA, Sewell WF. A possible neurotransmitter role for CGRP in a hair-cell sensory organ. Brain Res. 1987;419:347–351. - PubMed
-
- Altschuler RA, Parakkal MH, Fex J. Localization of enkephalin-like immunoreactivity in acetylcholinesterase-positive cells in the guinea-pig lateral superior olivary complex that project to the cochlea. Neuroscience. 1983;9:621–630. - PubMed
-
- Altschuler RA, Fex J, Parakkal MH, Eckenstein F. Colocalization of enkephalin-like and choline acetyltransferase-like immunoreactivities in olivocochlear neurons of the guinea pig. J. Histochem. Cytochem. 1984;32:839–843. - PubMed
-
- Altschuler RA, Hoffman DW, Reeks KA, Fex J. Localization of dynorphin B-like and alpha-neoendorphin-like immunoreactivities in the guinea pig organ of Corti. Hear. Res. 1985;17:249–258. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
