

Divalent cation sensitivity of BK channel activation supports the existence of three distinct binding sites
- PMID: 15738049
- PMCID: PMC2234011
- DOI: 10.1085/jgp.200409239
Divalent cation sensitivity of BK channel activation supports the existence of three distinct binding sites
Abstract
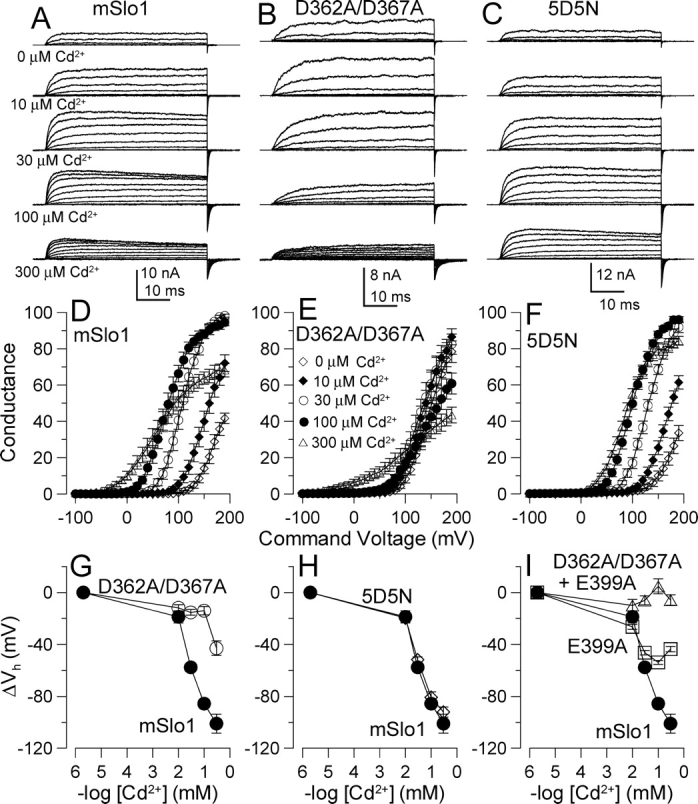
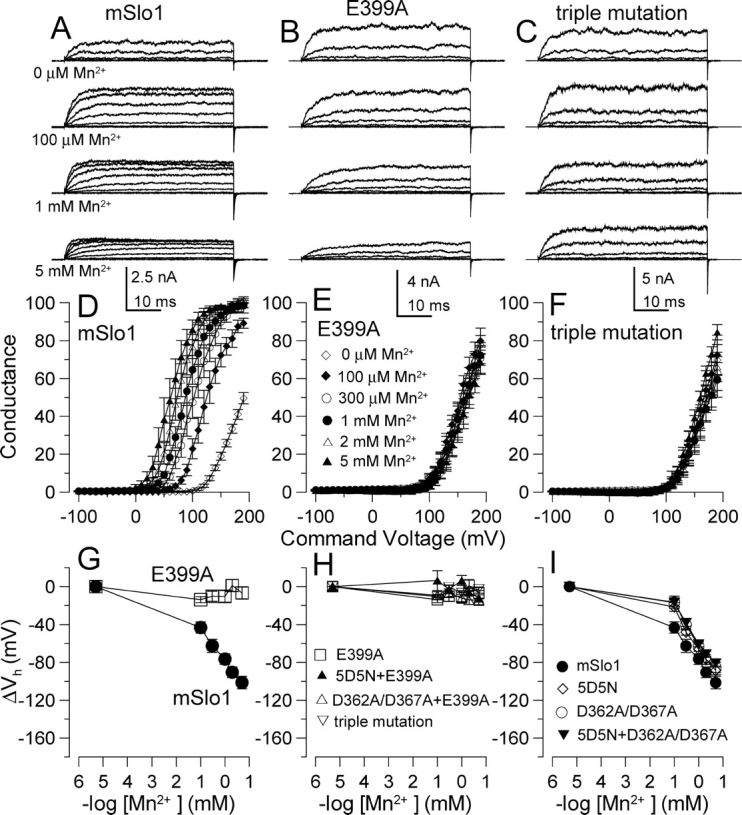
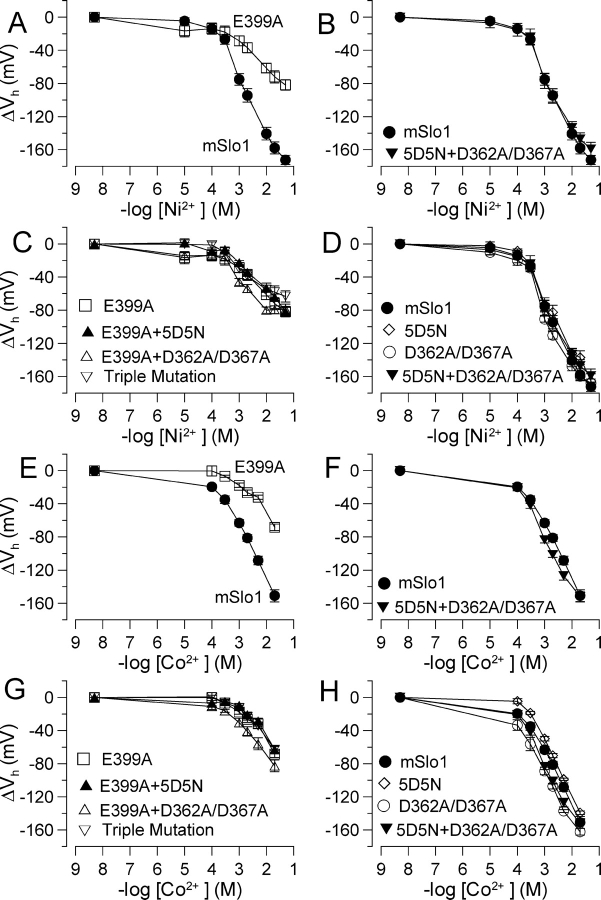
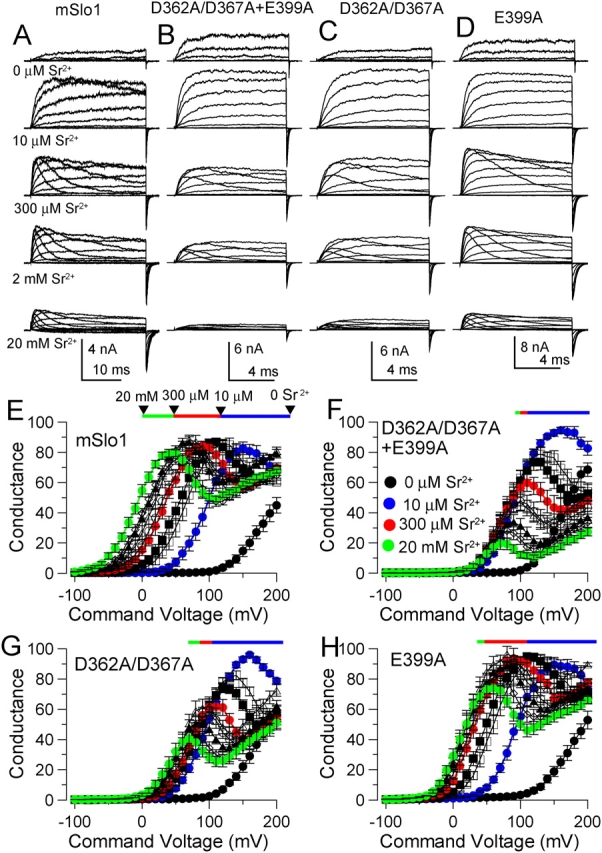
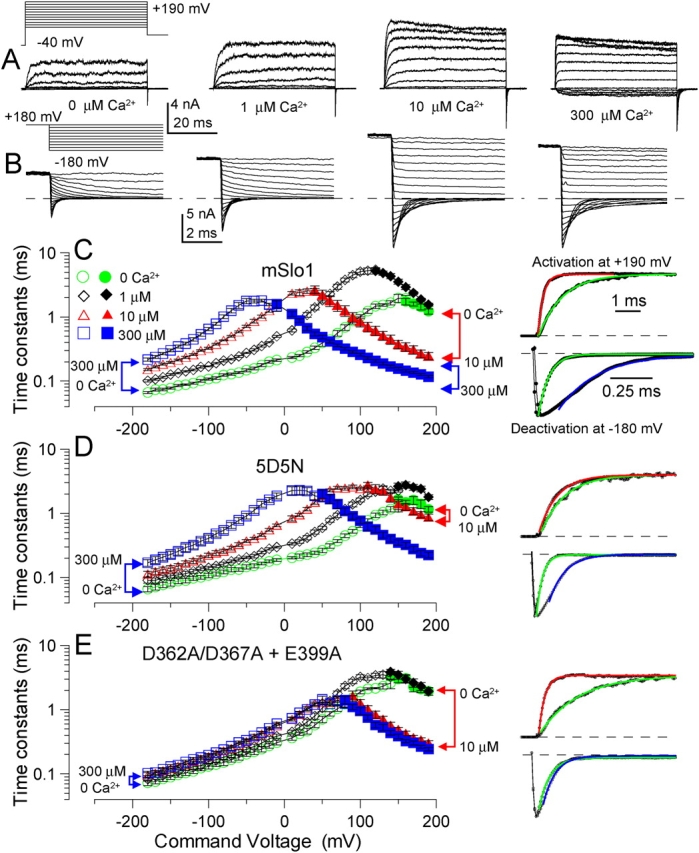
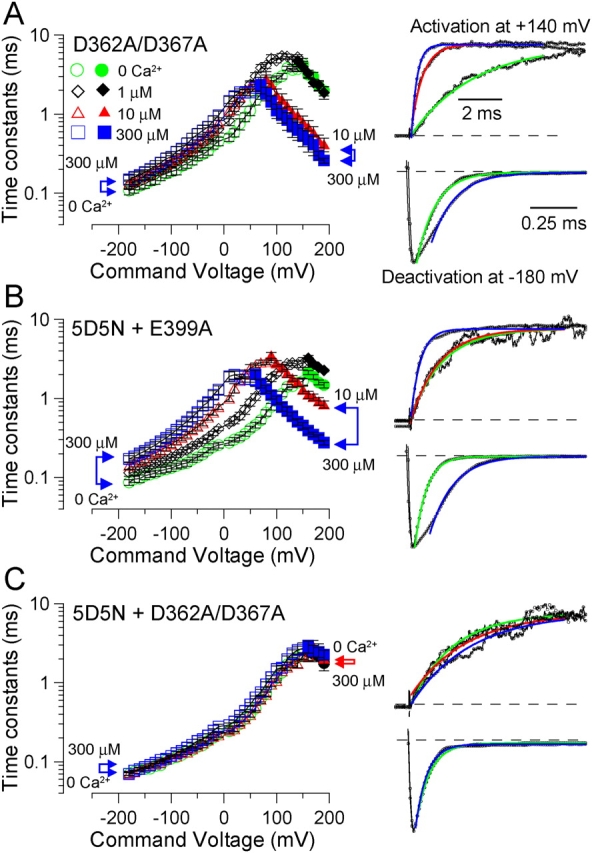
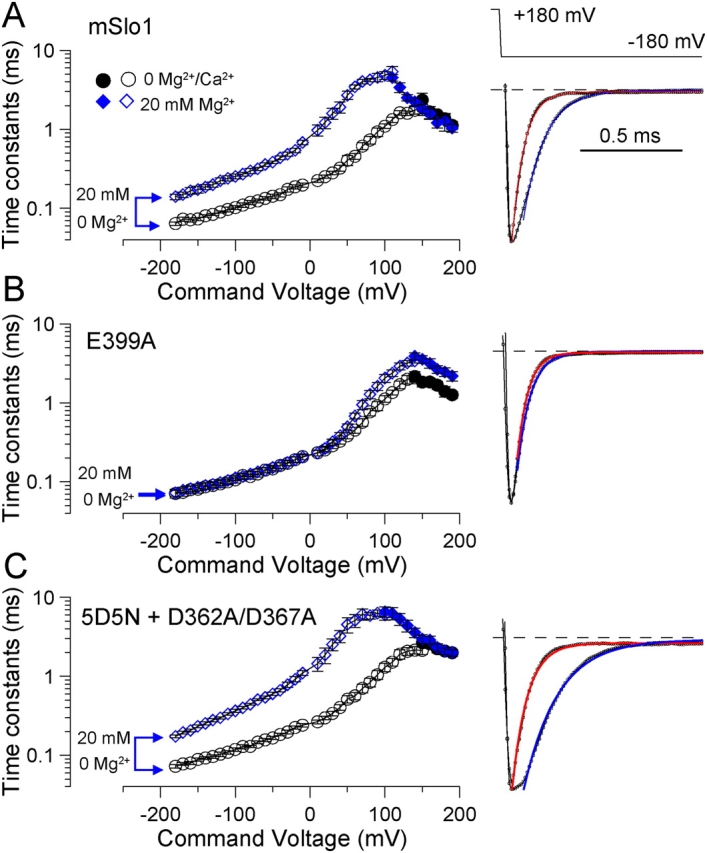
Mutational analyses have suggested that BK channels are regulated by three distinct divalent cation-dependent regulatory mechanisms arising from the cytosolic COOH terminus of the pore-forming alpha subunit. Two mechanisms account for physiological regulation of BK channels by microM Ca2+. The third may mediate physiological regulation by mM Mg2+. Mutation of five aspartate residues (5D5N) within the so-called Ca2+ bowl removes a portion of a higher affinity Ca2+ dependence, while mutation of D362A/D367A in the first RCK domain also removes some higher affinity Ca2+ dependence. Together, 5D5N and D362A/D367A remove all effects of Ca2+ up through 1 mM while E399A removes a portion of low affinity regulation by Ca2+/Mg2+. If each proposed regulatory effect involves a distinct divalent cation binding site, the divalent cation selectivity of the actual site that defines each mechanism might differ. By examination of the ability of various divalent cations to activate currents in constructs with mutationally altered regulatory mechanisms, here we show that each putative regulatory mechanism exhibits a unique sensitivity to divalent cations. Regulation mediated by the Ca2+ bowl can be activated by Ca2+ and Sr2+, while regulation defined by D362/D367 can be activated by Ca2+, Sr2+, and Cd2+. Mn2+, Co2+, and Ni2+ produce little observable effect through the high affinity regulatory mechanisms, while all six divalent cations enhance activation through the low affinity mechanism defined by residue E399. Furthermore, each type of mutation affects kinetic properties of BK channels in distinct ways. The Ca2+ bowl mainly accelerates activation of BK channels at low [Ca2+], while the D362/D367-related high affinity site influences both activation and deactivation over the range of 10-300 microM Ca2+. The major kinetic effect of the E399-related low affinity mechanism is to slow deactivation at mM Mg2+ or Ca2+. The results support the view that three distinct divalent-cation binding sites mediate regulation of BK channels.
Figures
Comment in
-
The BKCa channel's Ca2+-binding sites, multiple sites, multiple ions.J Gen Physiol. 2005 Mar;125(3):253-5. doi: 10.1085/jgp.200509270. J Gen Physiol. 2005. PMID: 15738047 Free PMC article. Review. No abstract available.
Similar articles
-
Multiple regulatory sites in large-conductance calcium-activated potassium channels.Nature. 2002 Aug 22;418(6900):880-4. doi: 10.1038/nature00956. Nature. 2002. PMID: 12192411
-
Beta1 subunits facilitate gating of BK channels by acting through the Ca2+, but not the Mg2+, activating mechanisms.Proc Natl Acad Sci U S A. 2003 Aug 19;100(17):10061-6. doi: 10.1073/pnas.1731650100. Epub 2003 Jul 31. Proc Natl Acad Sci U S A. 2003. PMID: 12893878 Free PMC article.
-
Interactions of divalent cations with calcium binding sites of BK channels reveal independent motions within the gating ring.Proc Natl Acad Sci U S A. 2016 Dec 6;113(49):14055-14060. doi: 10.1073/pnas.1611415113. Epub 2016 Nov 21. Proc Natl Acad Sci U S A. 2016. PMID: 27872281 Free PMC article.
-
Large conductance Ca2+-activated K+ (BK) channel: activation by Ca2+ and voltage.Biol Res. 2006;39(3):385-401. doi: 10.4067/s0716-97602006000300003. Epub 2006 Nov 7. Biol Res. 2006. PMID: 17106573 Review.
-
Cation channels in oocytes and early states of development: a novel type of nonselective cation channel activated by adrenaline in a clonal mesoderm-like cell line (MES-1).EXS. 1993;66:297-303. doi: 10.1007/978-3-0348-7327-7_24. EXS. 1993. PMID: 7505660 Review.
Cited by
-
Molecular mechanisms underlying the effect of the novel BK channel opener GoSlo: involvement of the S4/S5 linker and the S6 segment.Proc Natl Acad Sci U S A. 2015 Feb 17;112(7):2064-9. doi: 10.1073/pnas.1400555112. Epub 2015 Feb 4. Proc Natl Acad Sci U S A. 2015. PMID: 25653338 Free PMC article.
-
Barium ions selectively activate BK channels via the Ca2+-bowl site.Proc Natl Acad Sci U S A. 2012 Jul 10;109(28):11413-8. doi: 10.1073/pnas.1204444109. Epub 2012 Jun 25. Proc Natl Acad Sci U S A. 2012. PMID: 22733762 Free PMC article.
-
TRPC6 channels and their binding partners in podocytes: role in glomerular filtration and pathophysiology.Am J Physiol Renal Physiol. 2010 Oct;299(4):F689-701. doi: 10.1152/ajprenal.00298.2010. Epub 2010 Aug 4. Am J Physiol Renal Physiol. 2010. PMID: 20685822 Free PMC article. Review.
-
BK Channel Gating Mechanisms: Progresses Toward a Better Understanding of Variants Linked Neurological Diseases.Front Physiol. 2021 Oct 21;12:762175. doi: 10.3389/fphys.2021.762175. eCollection 2021. Front Physiol. 2021. PMID: 34744799 Free PMC article. Review.
-
State-dependent inhibition of BK channels by the opioid agonist loperamide.J Gen Physiol. 2021 Sep 6;153(9):e202012834. doi: 10.1085/jgp.202012834. Epub 2021 Aug 6. J Gen Physiol. 2021. PMID: 34357374 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
