

An immunoglobulin C kappa-reactive single chain antibody fusion protein induces tolerance through receptor editing in a normal polyclonal immune system
- PMID: 15738053
- PMCID: PMC2212821
- DOI: 10.1084/jem.20041854
An immunoglobulin C kappa-reactive single chain antibody fusion protein induces tolerance through receptor editing in a normal polyclonal immune system
Abstract
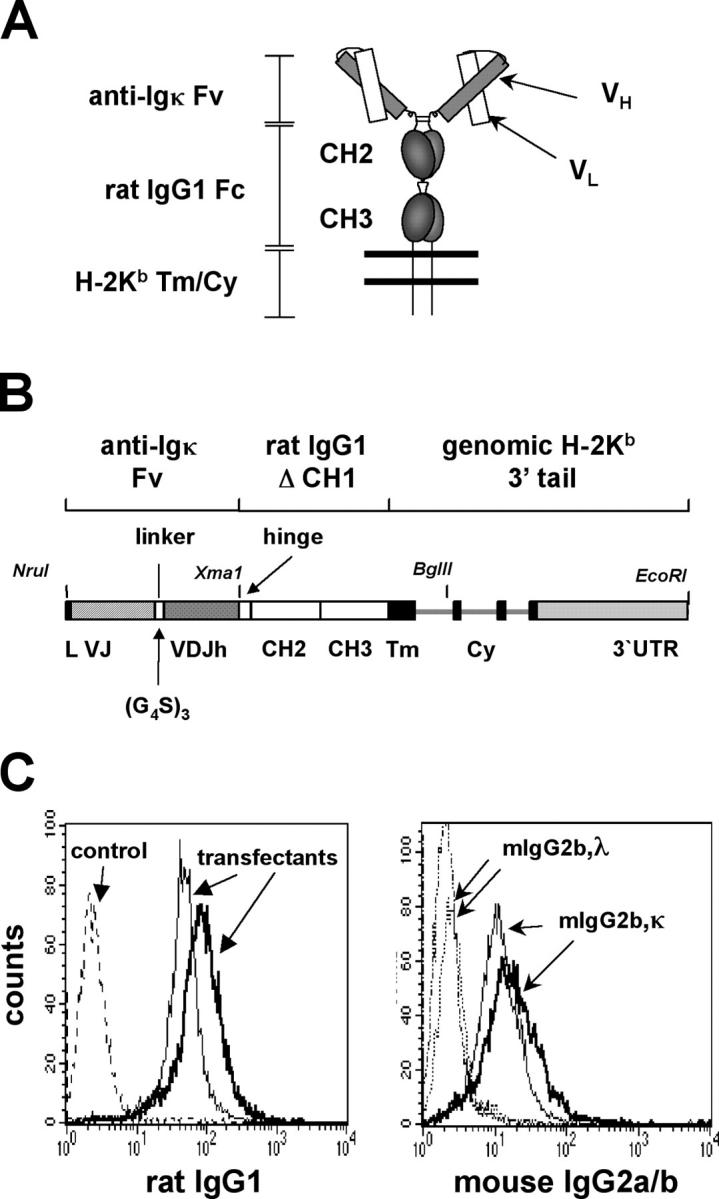
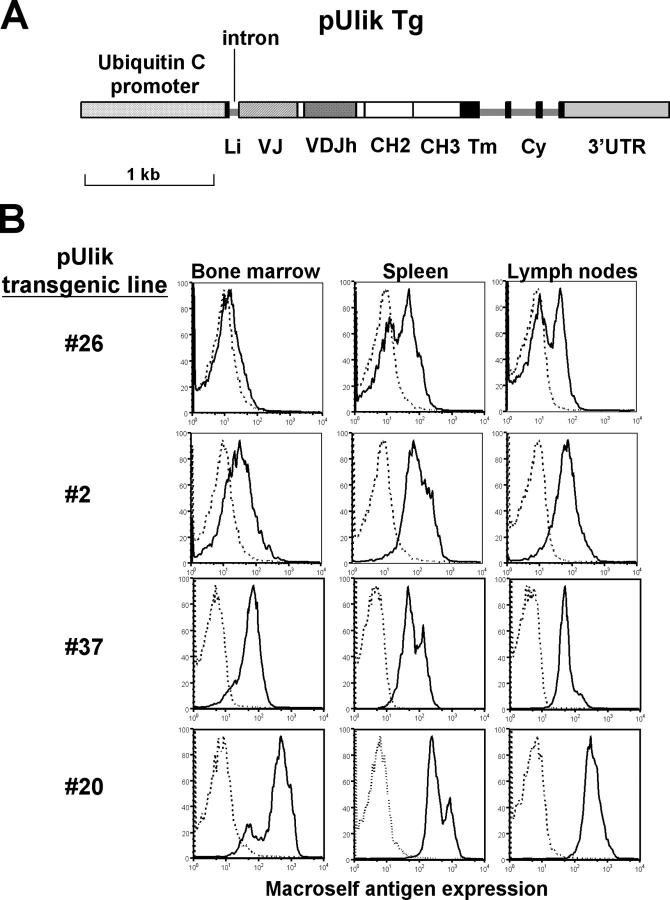
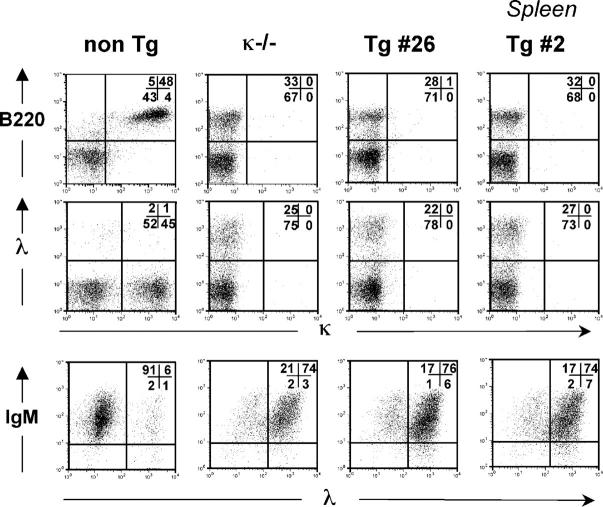
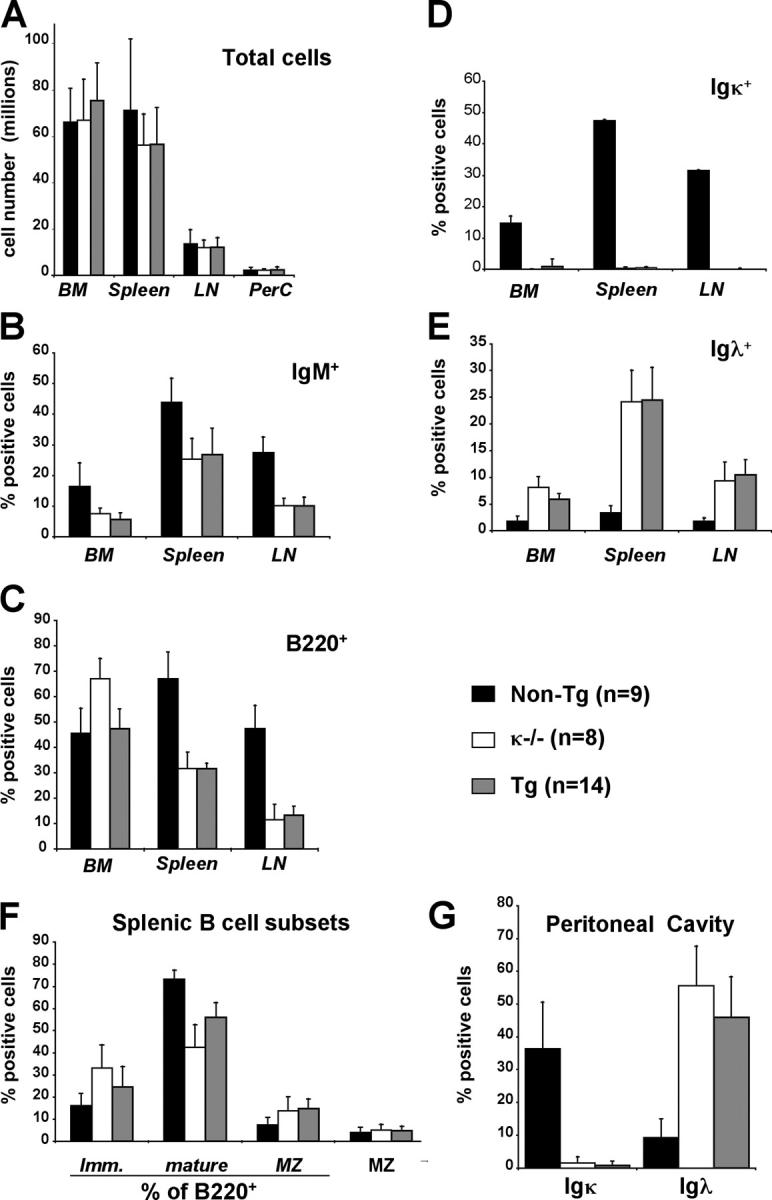
Understanding immune tolerance mechanisms is a major goal of immunology research, but mechanistic studies have generally required the use of mouse models carrying untargeted or targeted antigen receptor transgenes, which distort lymphocyte development and therefore preclude analysis of a truly normal immune system. Here we demonstrate an advance in in vivo analysis of immune tolerance that overcomes these shortcomings. We show that custom superantigens generated by single chain antibody technology permit the study of tolerance in a normal, polyclonal immune system. In the present study we generated a membrane-tethered anti-Igkappa-reactive single chain antibody chimeric gene and expressed it as a transgene in mice. B cell tolerance was directly characterized in the transgenic mice and in radiation bone marrow chimeras in which ligand-bearing mice served as recipients of nontransgenic cells. We find that the ubiquitously expressed, Igkappa-reactive ligand induces efficient B cell tolerance primarily or exclusively by receptor editing. We also demonstrate the unique advantages of our model in the genetic and cellular analysis of immune tolerance.
Figures
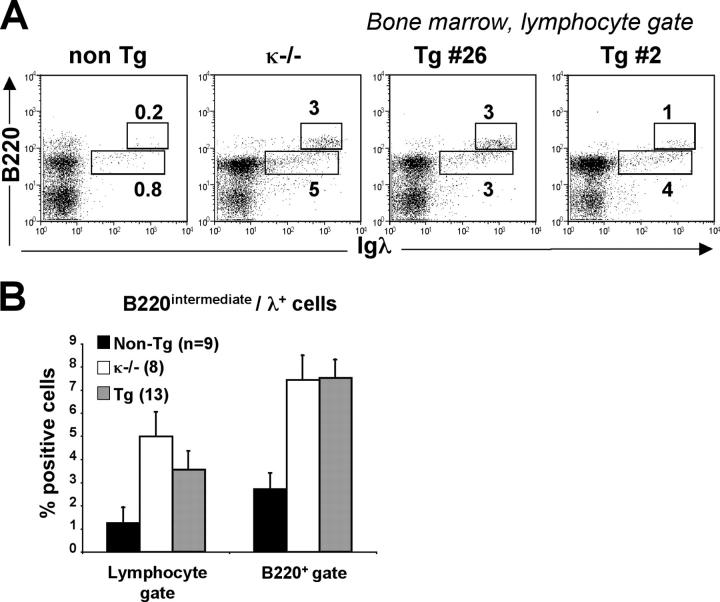
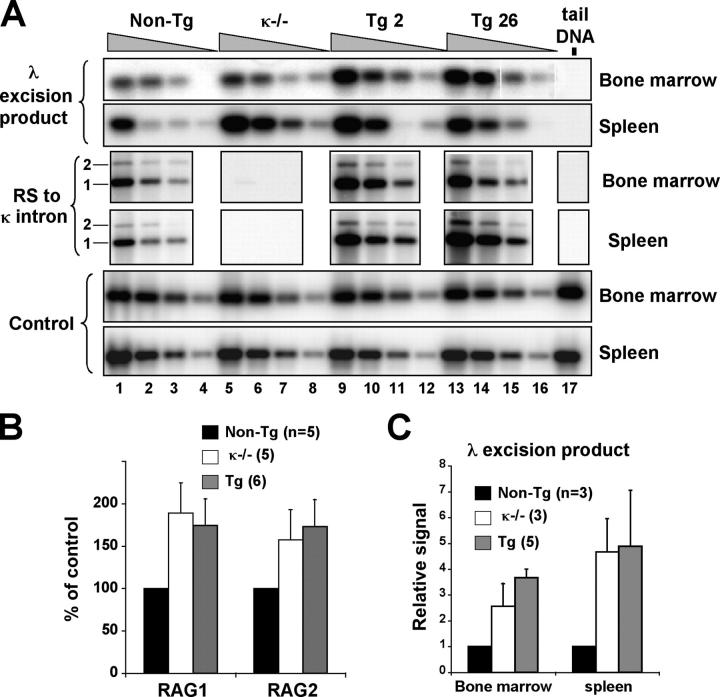
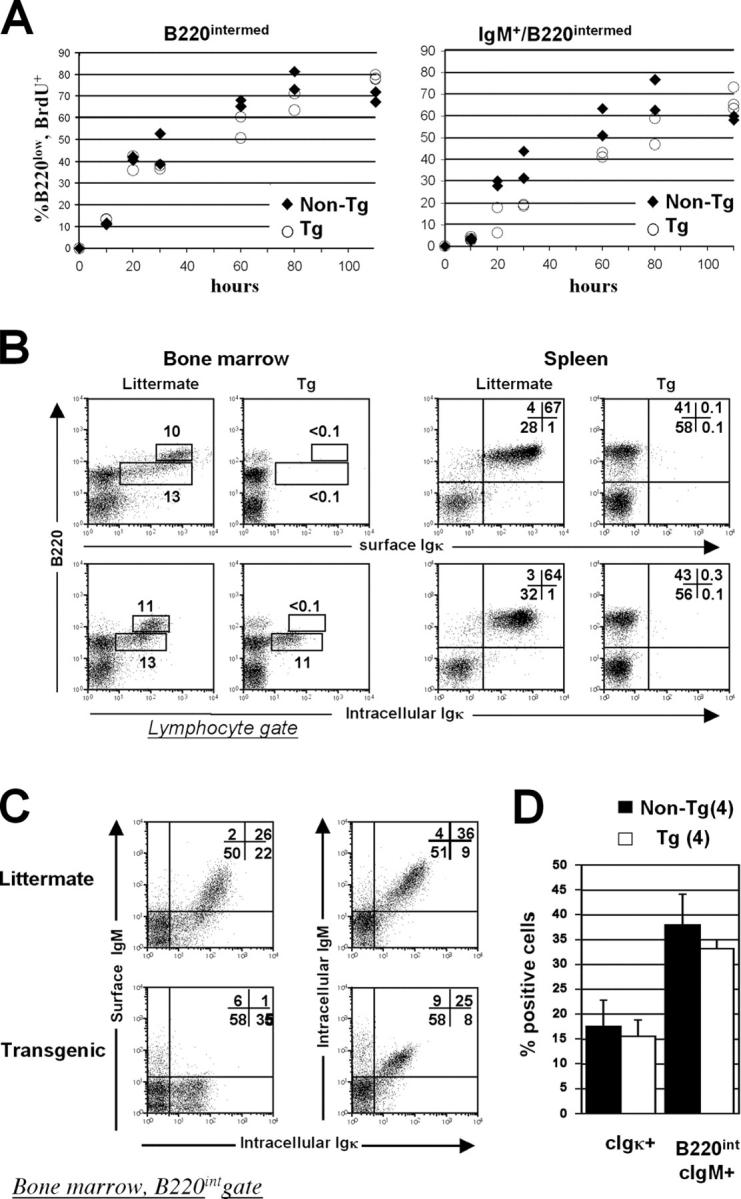
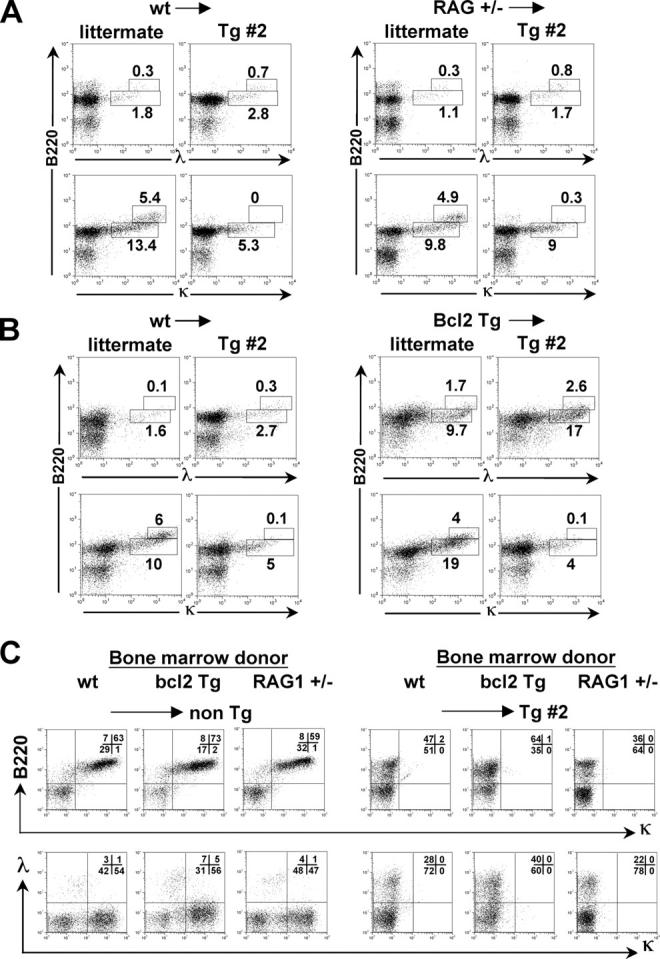
References
-
- Klinman, N.R. 1996. The “clonal selection hypothesis” and current concepts of B cell tolerance. Immunity. 5:189–195. - PubMed
-
- Nossal, G.J. 1983. Cellular mechanisms of immunologic tolerance. Annu. Rev. Immunol. 1:33–62. - PubMed
-
- Storb, U. 1987. Transgenic mice with immunoglobulin genes. Annu. Rev. Immunol. 5:151–174. - PubMed
-
- Goodnow, C.C. 1992. Transgenic mice and analysis of B-cell tolerance. Annu. Rev. Immunol. 10:489–518. - PubMed
-
- Miller, J.F., and A. Basten. 1996. Mechanisms of tolerance to self. Curr. Opin. Immunol. 8:815–821. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
