

Function and regulation of Tumbleweed (RacGAP50C) in neuroblast proliferation and neuronal morphogenesis
- PMID: 15738386
- PMCID: PMC553341
- DOI: 10.1073/pnas.0500748102
Function and regulation of Tumbleweed (RacGAP50C) in neuroblast proliferation and neuronal morphogenesis
Abstract
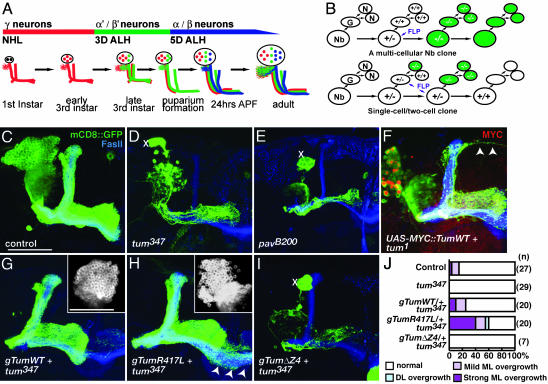
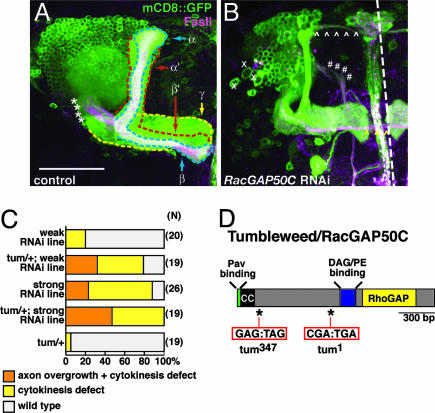
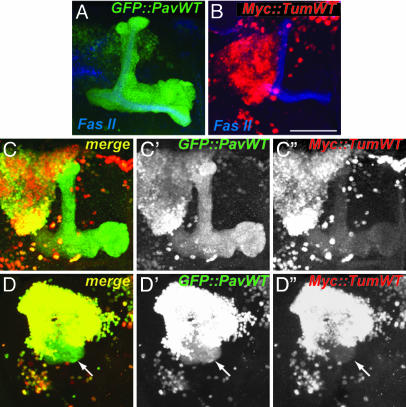
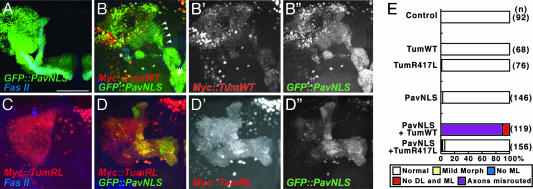
Drosophila RacGAP50C and its homologues act as part of a complex with a kinesin-like protein (Pavarotti/Zen-4) that is essential for the formation of the central spindle and completion of cytokinesis [Mishima, M., Kaitna, S. & Glotzer, M. (2002) Dev. Cell 2, 41-54; Somers, W. G. & Saint, R. (2003) Dev. Cell 4, 29-39; Jantsch-Plunger et al. (2000) J. Cell Biol. 149, 1391-1404]. We report here that RacGAP50C corresponds to the tumbleweed (tum) gene previously identified based on its defects in dendrite development of sensory neurons [Gao, F. B., Brenman, J. E., Jan, L. Y. & Jan, Y. N. (1999) Genes Dev. 13, 2549-2561]. Using mushroom body neurogenesis and morphogenesis as a model, we show that Tumbleweed (Tum), Pavarotti, and their association are required for neuroblast proliferation. Tum with a mutation predicted to disrupt the GTPase-activating protein (GAP) activity still largely retains its activity in regulating cell division but is impaired in its activity to limit axon growth. We also provide evidence that Tum and Pavarotti regulate the subcellular localization of each other in postmitotic neurons and that cytoplasmic accumulation of both proteins disrupts axon development in a GAP-dependent manner. Taken together with previous studies of RacGAP50C in regulating cytokinesis, we propose that Tum serves as a scaffolding protein in regulating cell division but acts as a GAP to limit axon growth in postmitotic neurons.
Figures
References
-
- Scheffzek, K., Ahmadian, M. R., Kabsch, W., Wiesmuller, L., Lautwein, A., Schmitz, F. & Wittinghofer, A. (1997) Science 277, 333-338. - PubMed
-
- Lander, E. S. (2001) Nature 409, 860-921. - PubMed
-
- Venter, J. C. (2001) Science 291, 1304-1351. - PubMed
-
- Billuart, P., Bienvenu, T., Ronce, N., des Portes, V., Vinet, M. C., Zemni, R., Roest Crollius, H., Carrie, A., Fauchereau, F., Cherry, M., et al. (1998) Nature 392, 923-926. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
