

Regulation of heterogenous nuclear ribonucleoprotein A1 transport by phosphorylation in cells stressed by osmotic shock
- PMID: 15738418
- PMCID: PMC553333
- DOI: 10.1073/pnas.0409889102
Regulation of heterogenous nuclear ribonucleoprotein A1 transport by phosphorylation in cells stressed by osmotic shock
Abstract
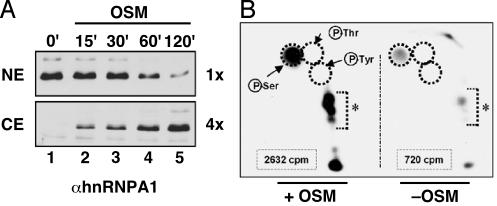
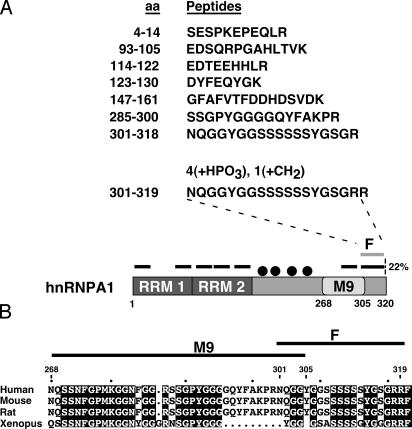
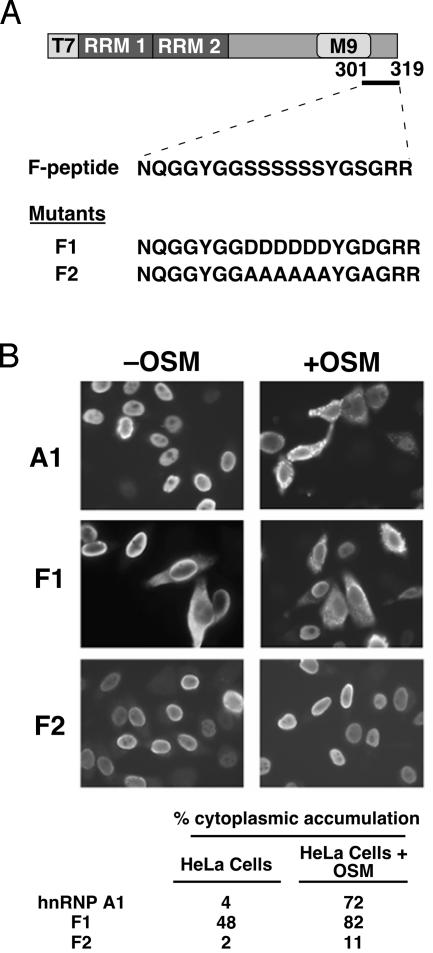
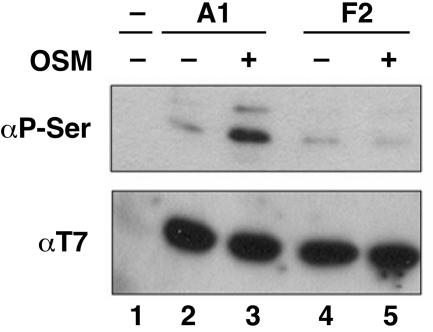
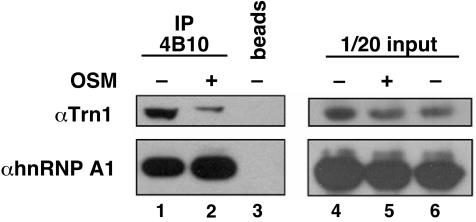
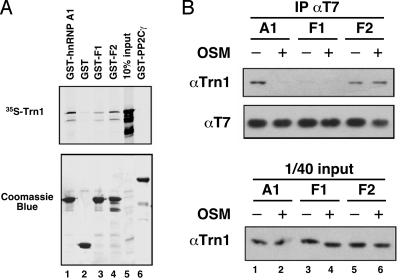
Heterogeneous nuclear ribonucleoprotein (hnRNP) A1 is an alternative splicing factor that is mainly nuclear, although it shuttles rapidly between nuclear and cytoplasmic compartments. Cells stressed by osmotic shock (OSM) activate the mitogen-activated protein kinase kinase(3/6)-p38 signaling pathway, which in turn results in accumulation of hnRNP A1 in the cytoplasm. This effect modulates alternative splicing regulation in vivo and correlates with increased hnRNP A1 phosphorylation. We have characterized the molecular mechanism involved in the cytoplasmic accumulation of hnRNP A1 in NIH 3T3 cells subjected to OSM. This treatment results in serine-specific phosphorylation within a C-terminal peptide, dubbed the "F-peptide," which is adjacent to the M9 motif that mediates bidirectional transport of hnRNP A1. Analysis of mutants in which the F-peptide serines were replaced by aspartic acids or alanines showed that F-peptide phosphorylation is required for the subcellular redistribution of hnRNP A1 in cells subjected to OSM. Furthermore, F-peptide phosphorylation modulates the interaction of hnRNP A1 with transportin Trn1. Our findings suggest that the phosphorylation of F-peptide by cell-signaling pathways regulates the rate of hnRNP A1 nuclear import.
Figures
References
-
- Dreyfuss, G., Kim, V. N. & Kataoka, N. (2002) Nat. Rev. Mol. Cell Biol. 3, 195-205. - PubMed
-
- Krecic, A. M. & Swanson, M. S. (1999) Curr. Opin. Cell Biol. 11, 363-371. - PubMed
-
- Piñol-Roma, S., Choi, Y. D., Matunis, M. J. & Dreyfuss, G. (1988) Genes Dev. 2, 215-227. - PubMed
-
- Dreyfuss, G., Matunis, M. J., Piñol-Roma, S. & Burd, C. G. (1993) Annu. Rev. Biochem. 62, 289-321. - PubMed
-
- Michael, W. M., Siomi, H., Choi, M., Piñol-Roma, S., Nakielny, S., Liu, Q. & Dreyfuss, G. (1995) Cold Spring Harb. Symp. Quant. Biol. 60, 663-668. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
