

doi: 10.1110/ps.041085005.
Epub 2005 Mar 1.
L-Arginine increases the solubility of unfolded species of hen egg white lysozyme
Affiliations
- PMID: 15741330
- PMCID: PMC2253432
- DOI: 10.1110/ps.041085005
Item in Clipboard
L-Arginine increases the solubility of unfolded species of hen egg white lysozyme
Protein Sci.
2005 Apr.
Abstract
L-Arginine (L-Arg) has been widely used as an enhancer of protein renaturation. The mechanism behind its action is still not fully understood. Using hen egg white lysozyme as a model protein, we present data that clearly demonstrate the suppression of the aggregation of denatured protein by L-Arg. By chemical modification of free cysteines, a series of unfolded lysozyme species were obtained that served as models for unfolded and intermediate states during the process of oxidative refolding. An increased equilibrium solubility of unfolded species and intermediates in the presence of L-Arg seems to be its major mechanism of action.
Figures
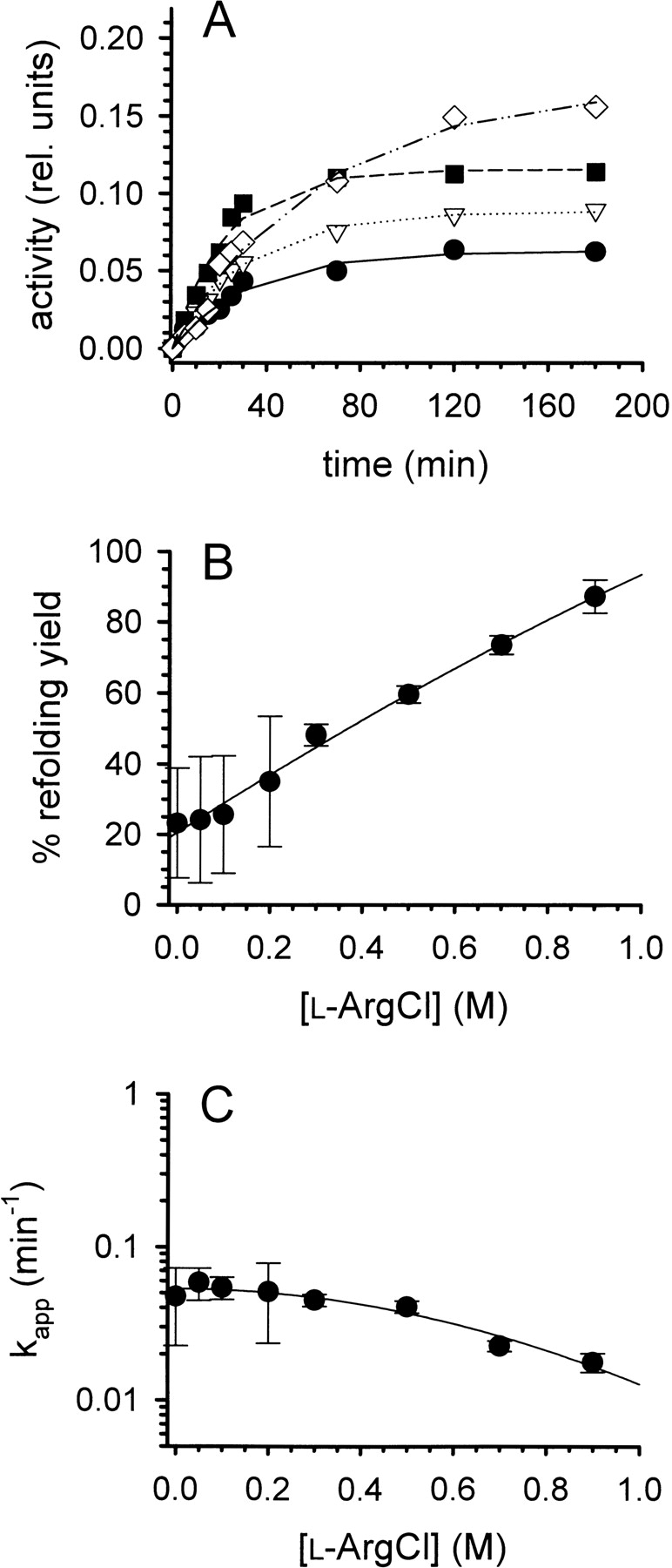
Refolding of lysozyme. (A) Denatured-reduced lysozyme was refolded at pH 8.2 and room temperature in the presence of a glutathione redox-shuffling system and increasing concentrations of l -ArgHCl as described in the text. The protein concentration was 0.25 mg mL−1. [l -ArgHCl] was 0, 0.2, 0.5, and 0.9 M (bottom to top). The refolding yield was monitored by measuring the enzymatic activity. (B) The final refolding yields were expressed as percentage of the specific activity of native lysozyme and plotted as function of [l -ArgHCl]. (C) The time course of refolding was analyzed as a pseudofirst-order reaction, yielding apparent time constants kapp. Data in B and C represent mean values ± SD from three independent experiments. Lines are meant to guide the eye.
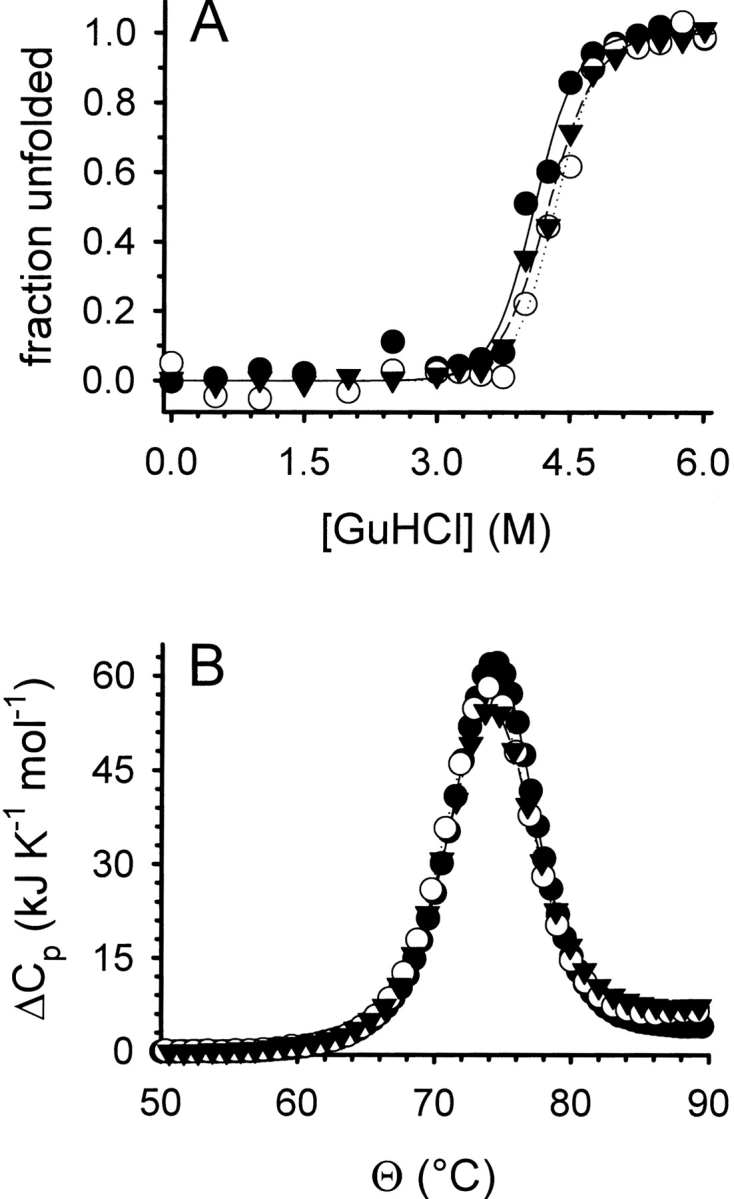
Denaturant-induced folding/unfolding (A) and thermal stability (B) of lysozyme. (A) The GuHCl-induced folding/unfolding equilibrium of lysozyme was monitored by tryptophan fluorescence at pH 7.0 without l -ArgHCl (filled circles), in the presence of 0.5 M l -ArgHCl (open circles), and of 1.0 M l -ArgHCl (filled triangles). Protein concentration was 20 μg mL−1. Data were normalized assuming a linear dependency of the intrinsic fluorescence for both the native and the denatured state. Lines represent fits to a two-state unfolding model. (B) The thermal unfolding of lysozyme was monitored by DSC at pH 6.0 without l -ArgHCl (filled circles), in the presence of 0.5 M l -ArgHCl (open circles), and of 1.0 M l -ArgHCl (filled triangles). Protein concentrations were 0.5 mg mL−1. Lines represent fits of the data to a two-state unfolding model.
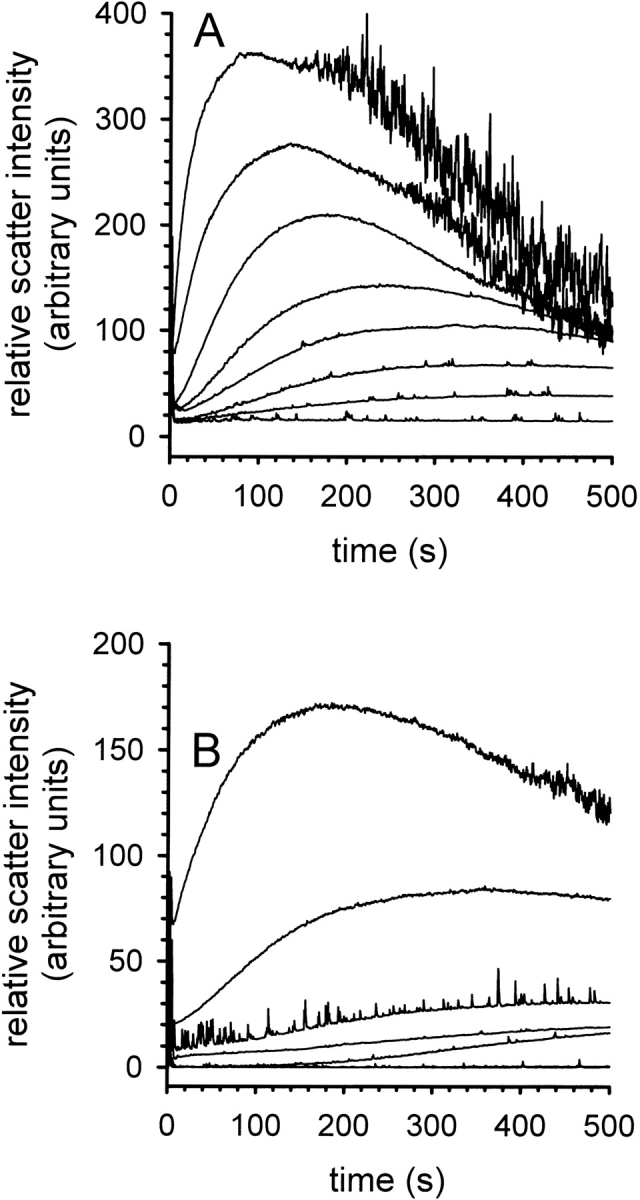
Kinetics of aggregation during the refolding of lysozyme. (A) Time-dependent formation of aggregates at increasing concentrations of denatured lysozyme in the presence of 50 mM l -ArgHCl. Lysozyme concentration was 0.1, 0.2, 0.25, 0.3, 0.35, 0.4, 0.5, and 0.6 mg mL−1 (bottom to top). (B) Time-dependent formation of aggregates in the presence of increasing concentrations of l -ArgHCl at a protein concentration of 0.3 mg mL−1. [l -ArgHCl] was 0, 0.05, 0.1, 0.15, 0.2, 0.25, and 0.4 M (top to bottom). The aggregation was monitored by light scattering at 600 nm.
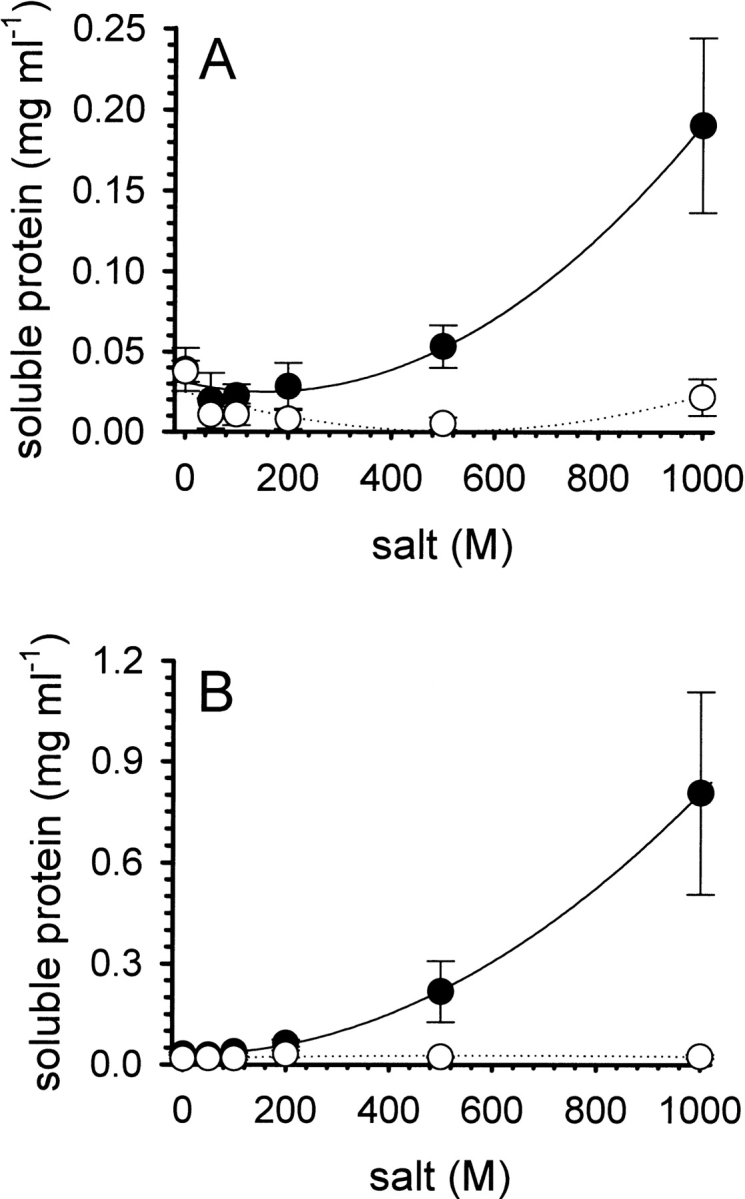
Solubility of iodoacetamide-modified (A) and iodoacetic acid-modified (B) denatured lysozyme. Preparations of the modified forms of denatured lysozyme (protein concentration ∼5 mg mL−1) were dialyzed overnight at room temperature against buffer at pH 8.5, containing increasing amounts of l -ArgHCl (filled circles) or NaCl (open circles). The concentrations of soluble protein in the supernatant were determined after centrifugation, as described in the text. Data represent mean values ± SD from four independent experiments. Lines are meant to guide the eye.
Similar articles
-
L-argininamide improves the refolding more effectively than L-arginine.J Biotechnol. 2007 Jun 15;130(2):153-60. doi: 10.1016/j.jbiotec.2007.03.003. Epub 2007 Mar 12. J Biotechnol. 2007. PMID: 17434637
-
Equilibrium and kinetic folding of hen egg-white lysozyme under acidic conditions.Proteins. 2002 Dec 1;49(4):472-82. doi: 10.1002/prot.10215. Proteins. 2002. PMID: 12402357
-
Oxidative refolding of reduced, denatured lysozyme in AOT reverse micelles.J Colloid Interface Sci. 2008 Jun 1;322(1):95-103. doi: 10.1016/j.jcis.2008.02.057. Epub 2008 Mar 4. J Colloid Interface Sci. 2008. PMID: 18377920
-
Protein crystallization under high pressure.Biochim Biophys Acta. 2002 Mar 25;1595(1-2):345-56. doi: 10.1016/s0167-4838(01)00355-7. Biochim Biophys Acta. 2002. PMID: 11983407 Review.
-
Review: Why is arginine effective in suppressing aggregation?Protein Pept Lett. 2005 Oct;12(7):613-9. doi: 10.2174/0929866054696109. Protein Pept Lett. 2005. PMID: 16522173 Review.
Cited by
-
A simplified method for the efficient purification and refolding of recombinant human TRAIL.Biotechnol Prog. 2020 Sep;36(5):e3007. doi: 10.1002/btpr.3007. Epub 2020 May 11. Biotechnol Prog. 2020. PMID: 32329219 Free PMC article.
-
Studying the mechanism of phase separation in aqueous solutions of globular proteins via molecular dynamics computer simulations.Phys Chem Chem Phys. 2021 Jan 6;23(1):415-424. doi: 10.1039/d0cp05160h. Phys Chem Chem Phys. 2021. PMID: 33319872 Free PMC article.
-
Effects of solutes on solubilization and refolding of proteins from inclusion bodies with high hydrostatic pressure.Protein Sci. 2006 Feb;15(2):304-13. doi: 10.1110/ps.051813506. Epub 2005 Dec 29. Protein Sci. 2006. PMID: 16385003 Free PMC article.
-
Quantification of anti-aggregation activity of chaperones: a test-system based on dithiothreitol-induced aggregation of bovine serum albumin.PLoS One. 2013 Sep 10;8(9):e74367. doi: 10.1371/journal.pone.0074367. eCollection 2013. PLoS One. 2013. PMID: 24058554 Free PMC article.
-
A change in the aggregation pathway of bovine serum albumin in the presence of arginine and its derivatives.Sci Rep. 2017 Jun 21;7(1):3984. doi: 10.1038/s41598-017-04409-x. Sci Rep. 2017. PMID: 28638090 Free PMC article.
References
-
- Acharya, A.S. and Taniuchi, H. 1982. Implication of the structure and stability of disulfide intermediates of lysozyme on the mechanism of renaturation. Mol. Cell. Biochem. 44 129–148. - PubMed
-
- Arakawa, T. and Tsumoto, K. 2003. The effects of arginine on refolding of aggregated proteins: Not facilitate refolding, but suppress aggregation. Biochem. Biophys. Res. Commun. 304 148–152. - PubMed
-
- Asano, R., Kudo, T., Makabe, K., Tsumoto, K., and Kumagai, I. 2002. Antitumor activity of interleukin-21 prepared by novel refolding procedure from inclusion bodies expressed in Escherichia coli. FEBS Lett. 528 70–76. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
