

Thiol-disulphide interchange in tubulin: kinetics and the effect on polymerization
- PMID: 15743274
- PMCID: PMC1175133
- DOI: 10.1042/BJ20042118
Thiol-disulphide interchange in tubulin: kinetics and the effect on polymerization
Abstract
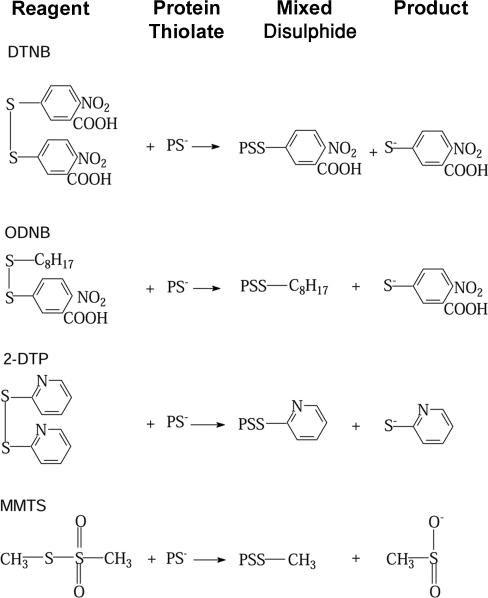
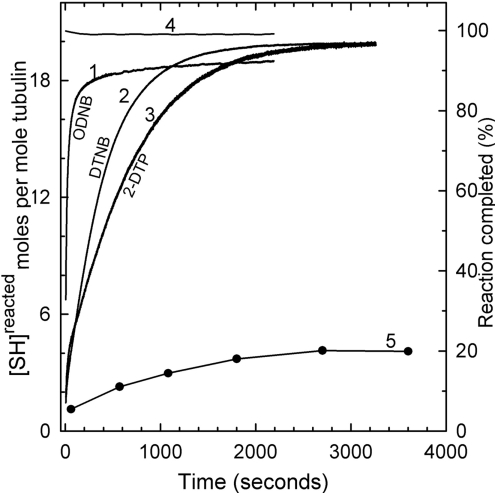
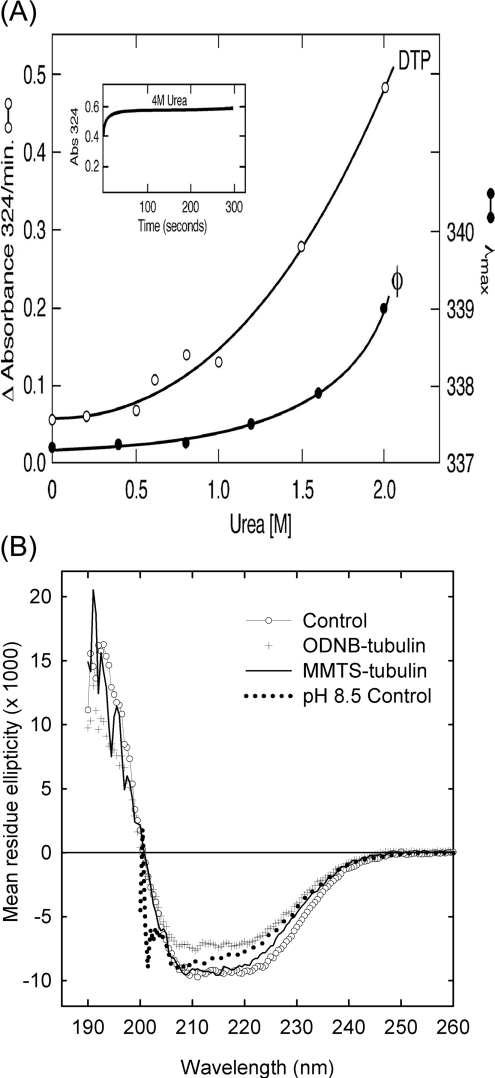
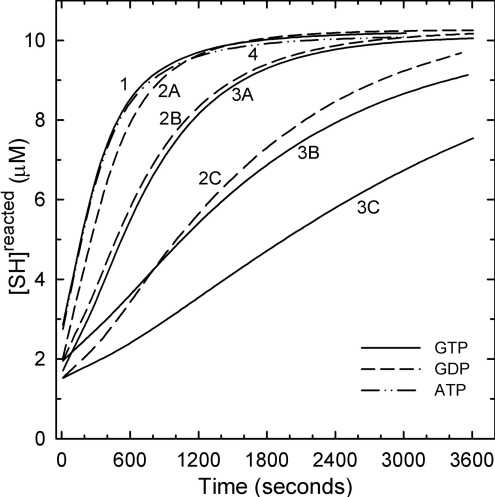
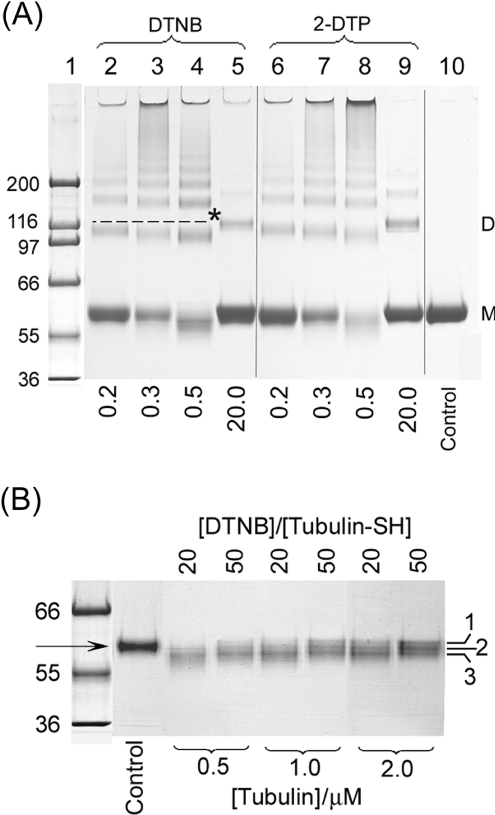
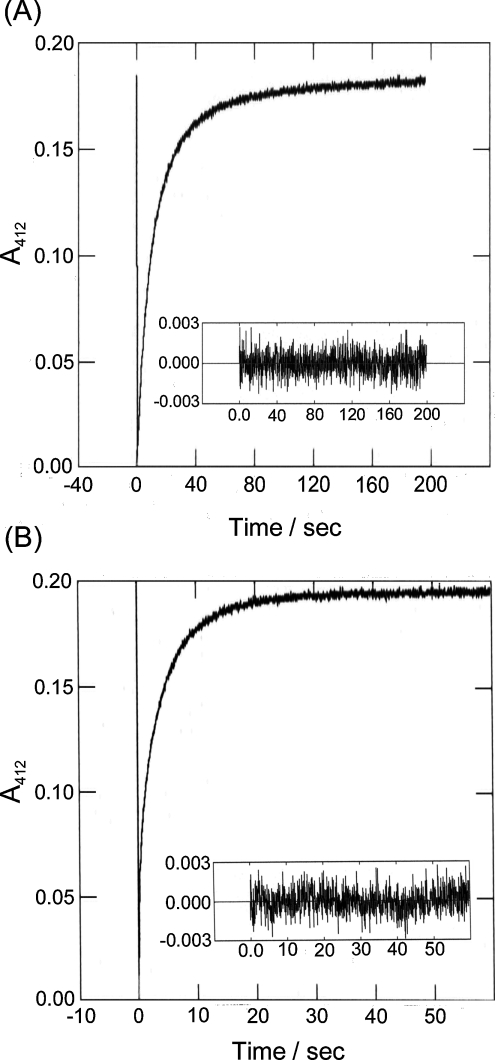
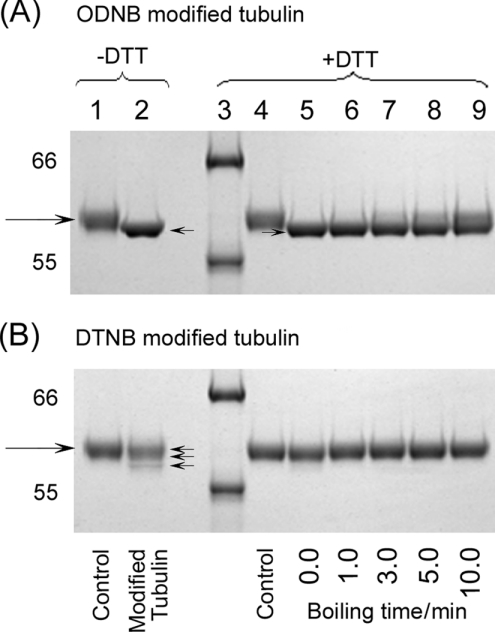
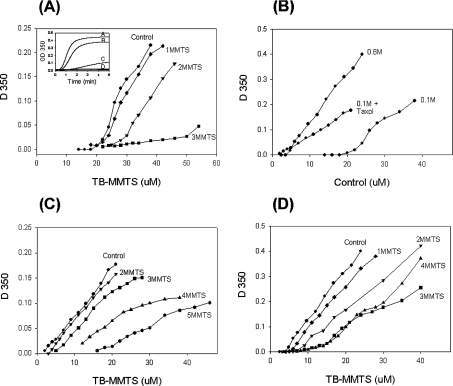
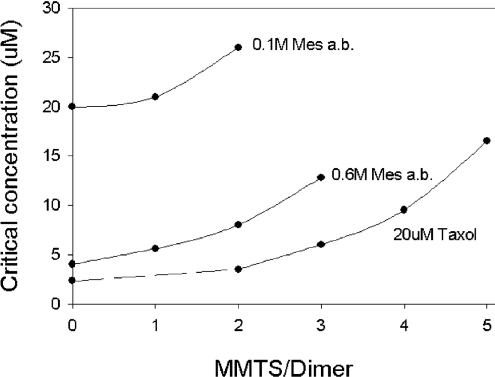
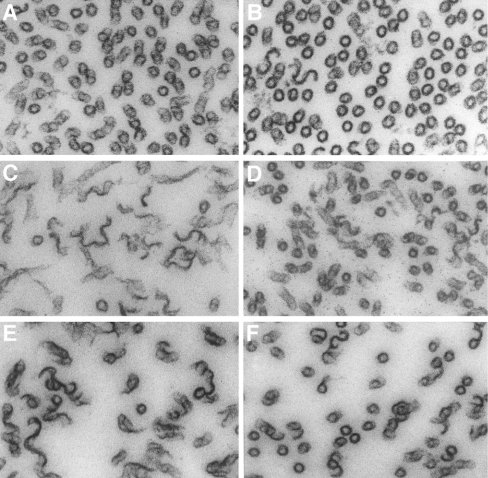
All 20 cysteine residues are accessible to disulphide reagents in the tubulin dimer, whereas only four are accessible in taxol-stabilized microtubules. Reaction rates with disulphide reagents are a function of the reagent, are decreased by G nucleotides, and increased with increase in pH and urea. With transient (stop-flow) kinetics, DTNB [5,5'-dithiobis-(2-nitrobenzoic acid)] and 2,2'-dithiodipyridine progress curves cannot be fitted by the sum of exponential terms based only on classes of cysteines. The mixed disulphide products react further to form both intra- and intermonomer disulphide bonds that can be reversed by reducing agents. With MMTS (methyl methanethiosulphonate) or ODNB (n-octyl-dithio-2-nitrobenzoate), virtually no protein-protein disulphide bonds are formed and the ODNB reaction can be given as the sum of three exponential terms with pseudo-first-order rate constants of 0.206, 0.069 and 0.010 s(-1) at pH 6.5, suggesting three classes of thiol reactivities. Limited cysteine substitution leads to only small changes in tryptophan or CD spectra, whereas complete substitution leads to loss of the helix content. MMTS-induced loss of SH groups leads to progressive increases in the critical concentration and loss of polymerization competence that can be reversed by assembly promoters such as higher protein concentration, taxol or high ionic strength. Under such conditions, the substituted tubulin forms protofilament-based structures such as microtubules, open tubules, sheets and/or bundles.
Figures
Similar articles
-
Exploration of the polar microenvironment around the reactive cysteine in rabbit muscle creatine kinase.Int J Biol Macromol. 2007 Oct 1;41(4):361-8. doi: 10.1016/j.ijbiomac.2007.05.004. Epub 2007 May 22. Int J Biol Macromol. 2007. PMID: 17592740
-
The SH-SS exchange reaction between the Ellman's reagent and protein containing SH groups as a method for determining conformational states: tubulin.Ital J Biochem. 1989 Mar-Apr;38(2):83-90. Ital J Biochem. 1989. PMID: 2745039
-
Determination and derivatization of protein thiols by n-octyldithionitrobenzoic acid.Anal Biochem. 1993 Feb 1;208(2):357-62. doi: 10.1006/abio.1993.1061. Anal Biochem. 1993. PMID: 8452233
-
Protein thiol modification by peroxynitrite anion and nitric oxide donors.Methods Enzymol. 2008;440:95-109. doi: 10.1016/S0076-6879(07)00805-1. Methods Enzymol. 2008. PMID: 18423212 Review.
-
A unique mechanism for thiolation of serum albumins by disulphide molecules.J Biochem. 2020 Feb 1;167(2):165-171. doi: 10.1093/jb/mvz084. J Biochem. 2020. PMID: 31598674 Review.
Cited by
-
Smokeless tobacco extract (STE)-induced toxicity in mammalian cells is mediated by the disruption of cellular microtubule network: a key mechanism of cytotoxicity.PLoS One. 2013 Jul 11;8(7):e68224. doi: 10.1371/journal.pone.0068224. Print 2013. PLoS One. 2013. PMID: 23874548 Free PMC article.
-
Inhibition of tubulin polymerization by hypochlorous acid and chloramines.Free Radic Biol Med. 2011 Apr 15;50(8):1000-8. doi: 10.1016/j.freeradbiomed.2011.01.018. Epub 2011 Jan 21. Free Radic Biol Med. 2011. PMID: 21256958 Free PMC article.
-
Molecular basis for class V beta-tubulin effects on microtubule assembly and paclitaxel resistance.J Biol Chem. 2009 May 8;284(19):13023-32. doi: 10.1074/jbc.M900167200. Epub 2009 Mar 12. J Biol Chem. 2009. PMID: 19282281 Free PMC article.
-
Proteomic analysis of covalent modifications of tubulins by isothiocyanates.J Nutr. 2012 Jul;142(7):1377S-81S. doi: 10.3945/jn.111.152041. Epub 2012 May 30. J Nutr. 2012. PMID: 22649267 Free PMC article.
-
The antioxidant protein alkylhydroperoxide reductase of Helicobacter pylori switches from a peroxide reductase to a molecular chaperone function.Proc Natl Acad Sci U S A. 2006 Feb 21;103(8):2552-7. doi: 10.1073/pnas.0510770103. Epub 2006 Feb 15. Proc Natl Acad Sci U S A. 2006. PMID: 16481626 Free PMC article.
References
-
- Cowan N. Tubulin genes and the diversity of microtubule function. Oxford Surveys on Eukaryotic Genes. In: Maclean N., editor. Oxford, U.K.: University of Oxford Press; 1984. pp. 26–60. - PubMed
-
- Lowe J., Li H., Downing K. H., Nogales E. Refined structure of alpha beta-tubulin at 3.5 A resolution. J. Mol. Biol. 2001;313:1045–1057. - PubMed
-
- Robinson A. S., King J. Disulphide-bonded intermediate on the folding and assembly pathway of a non-disulphide bonded protein. Nat. Struct. Biol. 1997;4:450–455. - PubMed
-
- Chaudhuri A. R., Khan I. A., Ludueña R. F. Detection of disulfide bonds in bovine brain tubulin and their role in protein folding and microtubule assembly in vitro: a novel disulfide detection approach. Biochemistry. 2001;40:8834–8841. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources