

CCR2-mediated recruitment of fibrocytes to the alveolar space after fibrotic injury
- PMID: 15743780
- PMCID: PMC1780139
- DOI: 10.1016/S0002-9440(10)62289-4
CCR2-mediated recruitment of fibrocytes to the alveolar space after fibrotic injury
Abstract
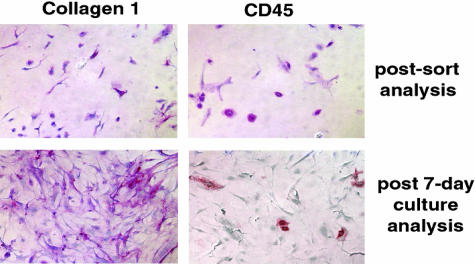
Bone marrow-derived cells are known to play important roles in repair/regeneration of injured tissues, but their roles in pathological fibrosis are less clear. Here, we report a critical role for the chemokine receptor CCR2 in the recruitment and activation of lung fibrocytes (CD45(+), CD13(+), collagen 1(+), CD34(-)). Lung fibrocytes were isolated in significantly greater numbers from airspaces of fluorescein isothiocyanate-injured CCR2(+/+) mice than from CCR2(-/-) mice. Transplant of CCR2(+/+) bone marrow into CCR2(-/-) recipients restored recruitment of lung fibrocytes and susceptibility to fibrosis. Ex vivo PKH-26-labeled CCR2(+/+) lung fibrocytes also migrated to injured airspaces of CCR2(-/-) recipients in vivo. Isolated lung fibrocytes expressed CCR2 and migrated to CCL2, and CCL2 stimulated collagen secretion by lung fibrocytes. Fibrocytes could transition into fibroblasts in vitro, and this transition was associated with loss of CCR2 expression and enhanced production of collagen 1. This is the first report describing expression of CCR2 on lung fibrocytes and demonstrating that CCR2 regulates both recruitment and activation of these cells after respiratory injury.
Figures







References
-
- Thannickal VJ, Toews GB, White E, Lynch JI, Martinez F. Mechanisms of pulmonary fibrosis. Annu Rev Med. 2004;55:395–471. - PubMed
-
- Lynch JI, Toews G: Idiopathic pulmonary fibrosis. Fishman A, Pulmonary Diseases and Disorders. Edited by Fishman A. Philadelphia PA, 1997, pp 1069-1084
-
- Kuhn C. Pathology. Phan S, Thrall R, editors. New York: Marcek Dekker, Inc.,; Pulmonary Fibrosis. 1995:pp 59–83.
-
- King TE, Jr, Schwarz MI, Brown K, Tooze JA, Colby TV, Waldron JA, Jr, Flint A, Thurlbeck W, Cherniack RM. Idiopathic pulmonary fibrosis: relationship between histopathologic features and mortality. Am J Respir Crit Care Med. 2001;164:1025–1032. - PubMed
-
- Coultas D, Zumwalt R, Black W, Sobonya R. The epidemiology of interstitial lung disease. Am J Respir Crit Care Med. 1994;150:967–972. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
