

Genetic and functional interaction of evolutionarily conserved regions of the Prp18 protein and the U5 snRNA
- PMID: 15743809
- PMCID: PMC1061626
- DOI: 10.1128/MCB.25.6.2107-2116.2005
Genetic and functional interaction of evolutionarily conserved regions of the Prp18 protein and the U5 snRNA
Abstract
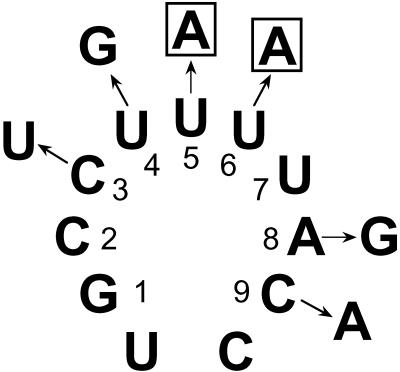
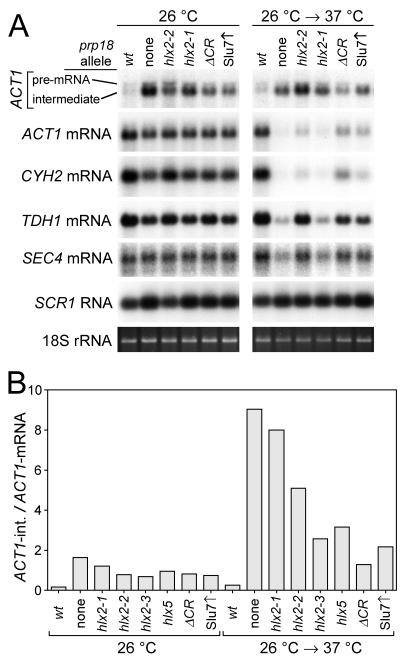
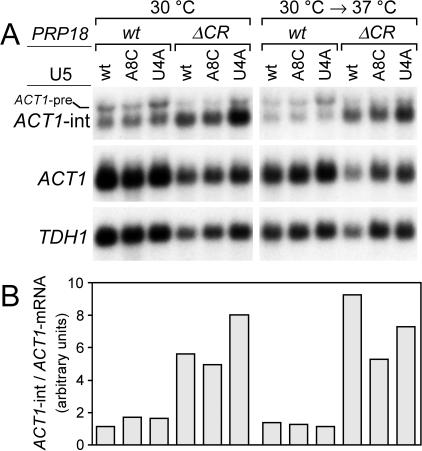
Both the Prp18 protein and the U5 snRNA function in the second step of pre-mRNA splicing. We identified suppressors of mutant prp18 alleles in the gene for the U5 snRNA (SNR7). The suppressors' U5 snRNAs have either a U4-to-A or an A8-to-C mutation in the evolutionarily invariant loop 1 of U5. Suppression is specific for prp18 alleles that encode proteins with mutations in a highly conserved region of Prp18 which forms an unstructured loop in crystals of Prp18. The snr7 suppressors partly restored the pre-mRNA splicing activity that was lost in the prp18 mutants. The close functional relationship of Prp18 and U5 is emphasized by the finding that two snr7 alleles, U5A and U6A, are dominant synthetic lethal with prp18 alleles. Our results support the idea that Prp18 and the U5 snRNA act in concert during the second step of pre-mRNA splicing and suggest a model in which the conserved loop of Prp18 acts to stabilize the interaction of loop 1 of the U5 snRNA with the splicing intermediates.
Figures
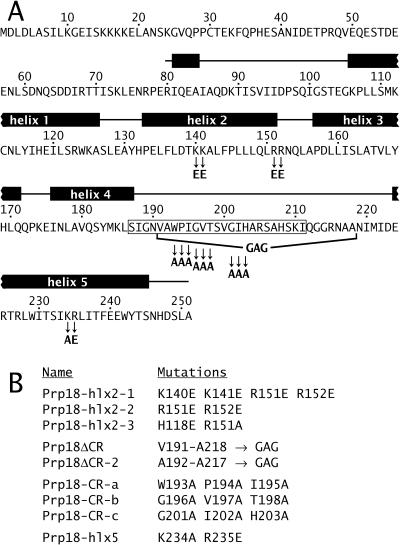
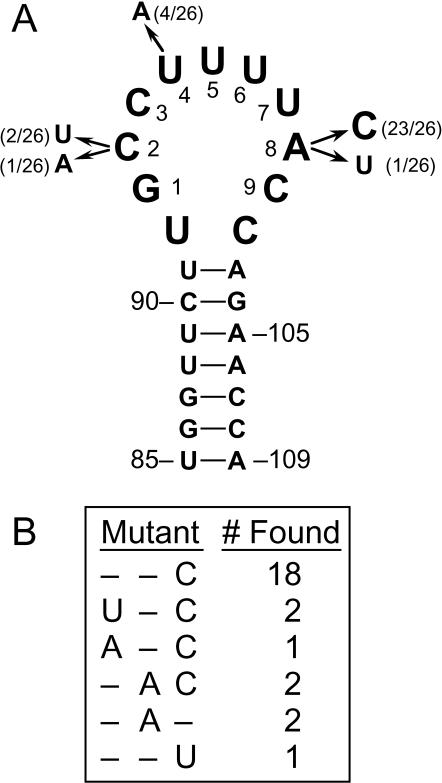
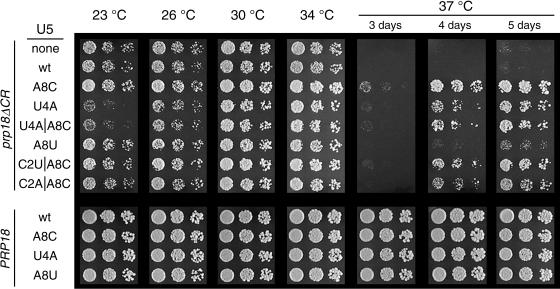
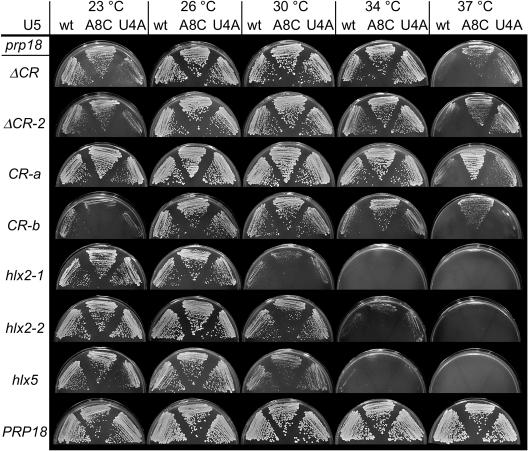

References
-
- Beggs, J. D., S. Teigelkamp, and A. J. Newman. 1995. The role of PRP8 protein in nuclear pre-mRNA splicing in yeast. J. Cell Sci. 19(Suppl.):101-105. - PubMed
-
- Bousquet-Antonelli, C., C. Presutti, and D. Tollervey. 2000. Identification of a regulated pathway for nuclear pre-mRNA turnover. Cell 102:765-775. - PubMed
-
- Brow, D. A. 2002. Allosteric cascade of spliceosome activation. Annu. Rev. Genet. 36:333-360. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases