

Inactivation of the Sema5a gene results in embryonic lethality and defective remodeling of the cranial vascular system
- PMID: 15743826
- PMCID: PMC1061610
- DOI: 10.1128/MCB.25.6.2310-2319.2005
Inactivation of the Sema5a gene results in embryonic lethality and defective remodeling of the cranial vascular system
Erratum in
-
Correction for Fiore et al., "Inactivation of the Sema5a Gene Results in Embryonic Lethality and Defective Remodeling of the Cranial Vascular System".Mol Cell Biol. 2020 Sep 28;40(20):e00409-20. doi: 10.1128/MCB.00409-20. Print 2020 Sep 28. Mol Cell Biol. 2020. PMID: 32989048 Free PMC article. No abstract available.
Abstract
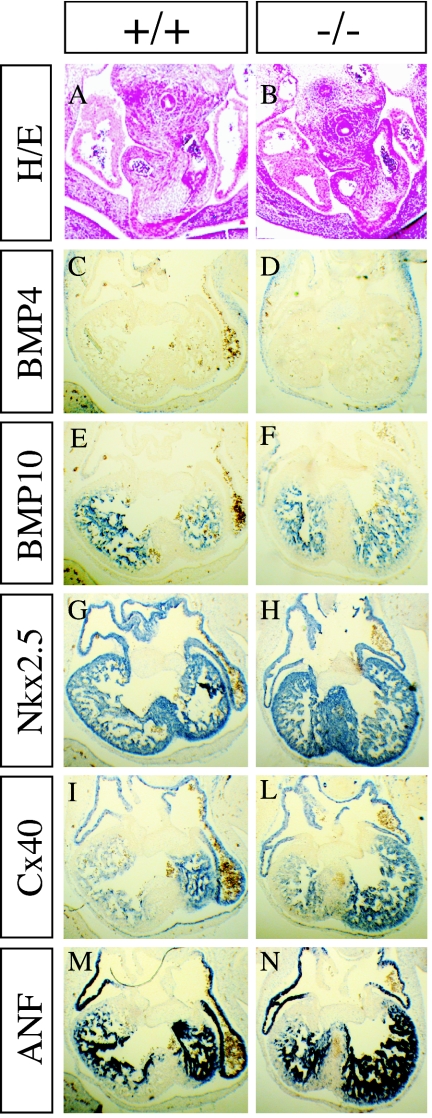
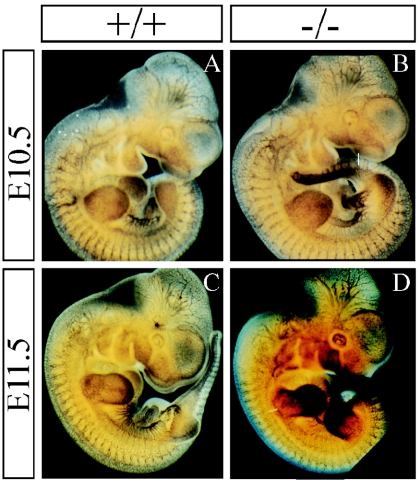
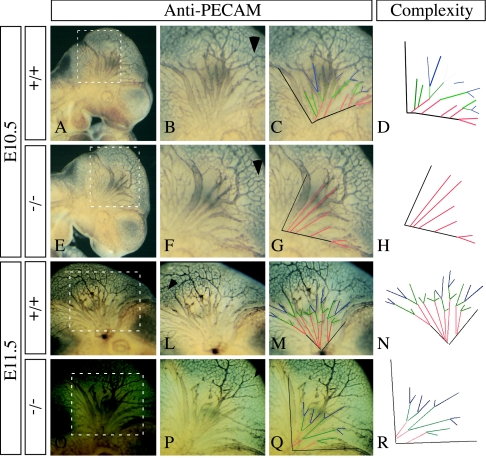
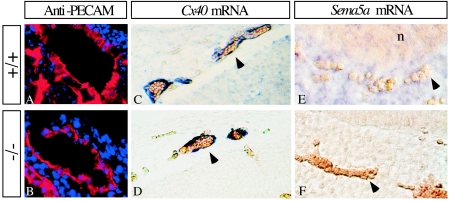
The semaphorins are a large family of proteins involved in the patterning of both the vascular and the nervous systems. In order to analyze the function of the membrane-bound semaphorin 5A (Sema5A), we generated mice homozygous for a null mutation in the Sema5a gene. Homozygous null mutants die between embryonic development days 11.5 (E11.5) and E12.5, indicating an essential role of Sema5A during embryonic development. Mutant embryos did not show any morphological defects that could account for the lethality of the mutation. A detailed analysis of the vascular system uncovered a role of Sema5A in the remodeling of the cranial blood vessels. In Sema5A null mutants, the complexity of the hierarchically organized branches of the cranial cardinal veins was decreased. Our results represent the first genetic analysis of the function of a class 5 semaphorin during embryonic development and identify a role of Sema5A in the regional patterning of the vasculature.
Figures
Similar articles
-
Semaphorin 5A is a bifunctional axon guidance cue for axial motoneurons in vivo.Dev Biol. 2009 Feb 1;326(1):190-200. doi: 10.1016/j.ydbio.2008.11.007. Epub 2008 Nov 20. Dev Biol. 2009. PMID: 19059233
-
Semaphorin 5A promotes angiogenesis by increasing endothelial cell proliferation, migration, and decreasing apoptosis.Microvasc Res. 2010 Jan;79(1):1-9. doi: 10.1016/j.mvr.2009.10.005. Epub 2009 Oct 19. Microvasc Res. 2010. PMID: 19850054 Free PMC article.
-
Multiple roles of mouse Numb in tuning developmental cell fates.Curr Biol. 2001 Apr 3;11(7):494-501. doi: 10.1016/s0960-9822(01)00149-x. Curr Biol. 2001. PMID: 11412999
-
Semaphorin function in the developing invertebrate peripheral nervous system.Biochem Cell Biol. 2000;78(5):603-11. Biochem Cell Biol. 2000. PMID: 11103951 Review.
-
Early embryonic lethality in genetically engineered mice: diagnosis and phenotypic analysis.Vet Pathol. 2012 Jan;49(1):64-70. doi: 10.1177/0300985810395725. Epub 2011 Jan 13. Vet Pathol. 2012. PMID: 21233329 Free PMC article. Review.
Cited by
-
Contribution of semaphorins to the formation of the peripheral nervous system in higher vertebrates.Cell Adh Migr. 2016 Nov;10(6):593-603. doi: 10.1080/19336918.2016.1243644. Epub 2016 Oct 7. Cell Adh Migr. 2016. PMID: 27715392 Free PMC article. Review.
-
Axon guidance and synaptic maintenance: preclinical markers for neurodegenerative disease and therapeutics.Trends Neurosci. 2009 Mar;32(3):142-9. doi: 10.1016/j.tins.2008.11.006. Epub 2009 Jan 21. Trends Neurosci. 2009. PMID: 19162339 Free PMC article. Review.
-
Semaphorins in Angiogenesis and Autoimmune Diseases: Therapeutic Targets?Front Immunol. 2020 Mar 5;11:346. doi: 10.3389/fimmu.2020.00346. eCollection 2020. Front Immunol. 2020. PMID: 32210960 Free PMC article. Review.
-
KIAA1199 as a potential diagnostic biomarker of rheumatoid arthritis related to angiogenesis.Arthritis Res Ther. 2015 May 29;17(1):140. doi: 10.1186/s13075-015-0637-y. Arthritis Res Ther. 2015. PMID: 26022278 Free PMC article.
-
Semaphorin signaling in cardiovascular development.Cell Metab. 2015 Feb 3;21(2):163-173. doi: 10.1016/j.cmet.2014.12.015. Cell Metab. 2015. PMID: 25651171 Free PMC article. Review.
References
-
- Adams, R. H., H. Betz, and A. W. Püschel. 1996. A novel class of murine semaphorins with homology to thrombospondin is differentially expressed during early embryogenesis. Mech. Dev. 57:33-45. - PubMed
-
- Adams, R. H., A. Porras, G. Alonso, M. Jones, K. Vintersten, S. Panelli, A. Valladares, L. Perez, R. Klein, and A. R. Nebreda. 2000. Essential role of p38alpha MAP kinase in placental but not embryonic cardiovascular development. Mol. Cell 6:109-116. - PubMed
-
- Adams, R. H., G. A. Wilkinson, C. Weiss, F. Diella, N. W. Gale, U. Deutsch, W. Risau, and R. Klein. 1999. Roles of ephrinB ligands and EphB receptors in cardiovascular development: demarcation of arterial/venous domains, vascular morphogenesis, and sprouting angiogenesis. Genes Dev. 13:295-306. - PMC - PubMed
-
- Bahri, S. M., W. Chia, and X. Yang. 2001. Characterization and mutant analysis of the Drosophila sema 5c gene. Dev. Dyn. 221:322-330. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases