

Neurabin/protein phosphatase-1 complex regulates dendritic spine morphogenesis and maturation
- PMID: 15743906
- PMCID: PMC1087240
- DOI: 10.1091/mbc.e04-12-1054
Neurabin/protein phosphatase-1 complex regulates dendritic spine morphogenesis and maturation
Abstract
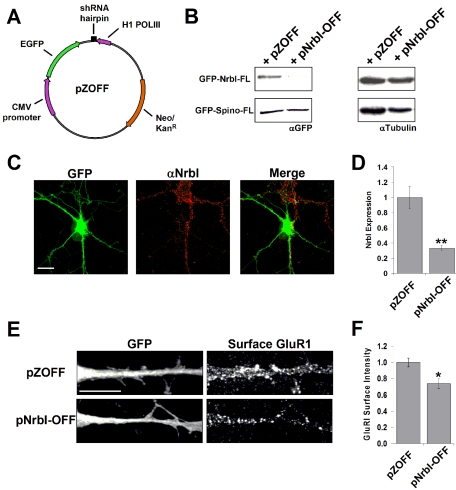
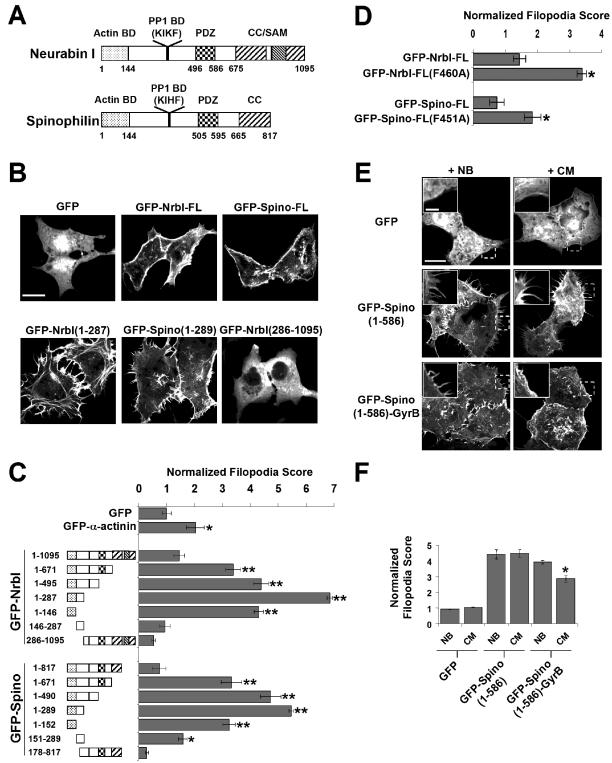
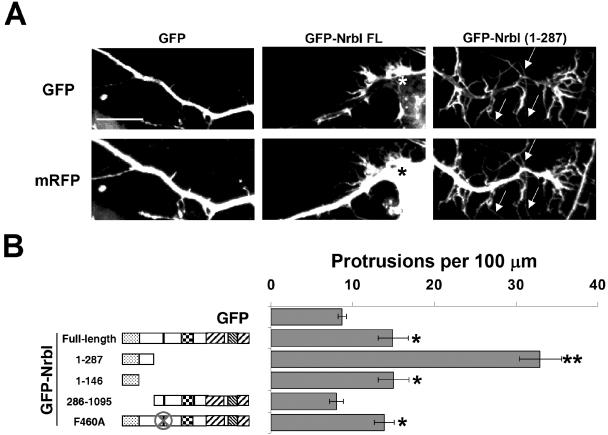
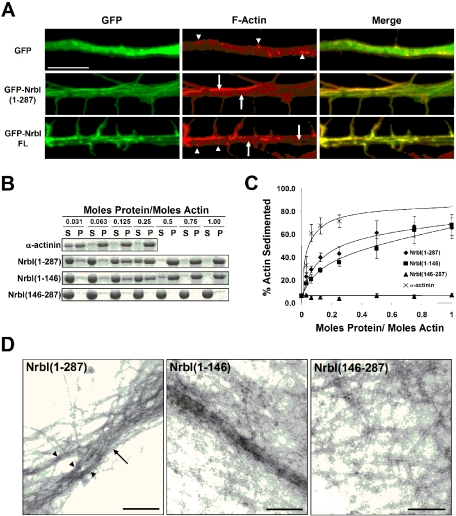
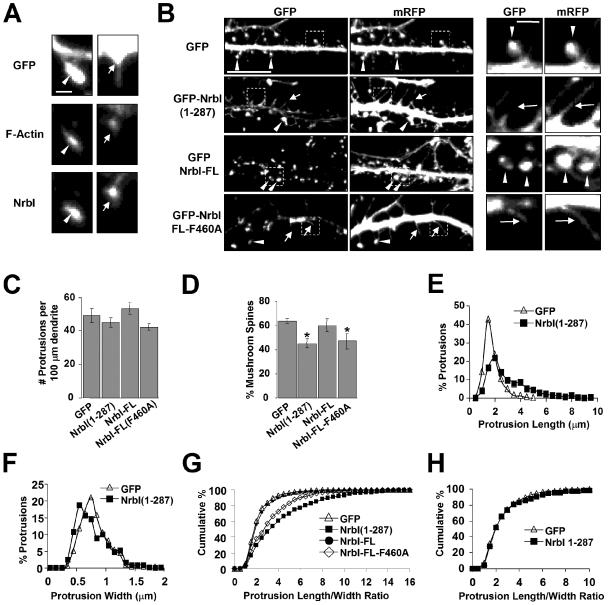
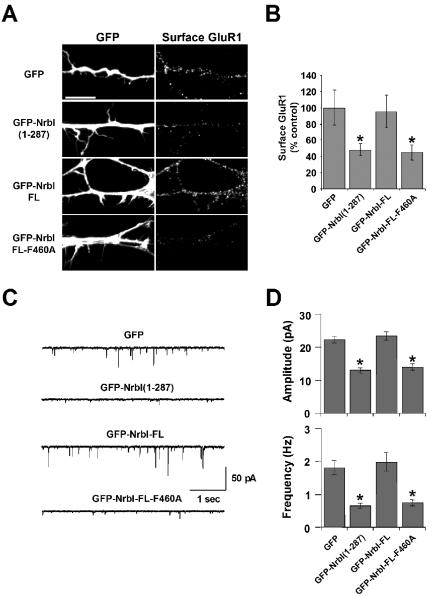
The majority of excitatory synapses in the mammalian brain form on filopodia and spines, actin-rich membrane protrusions present on neuronal dendrites. The biochemical events that induce filopodia and remodel these structures into dendritic spines remain poorly understood. Here, we show that the neuronal actin- and protein phosphatase-1-binding protein, neurabin-I, promotes filopodia in neurons and nonneuronal cells. Neurabin-I actin-binding domain bundled F-actin, promoted filopodia, and delayed the maturation of dendritic spines in cultured hippocampal neurons. In contrast, dimerization of neurabin-I via C-terminal coiled-coil domains and association of protein phosphatase-1 (PP1) with neurabin-I through a canonical KIXF motif inhibited filopodia. Furthermore, the expression of a neurabin-I polypeptide unable to bind PP1 delayed the maturation of neuronal filopodia into spines, reduced the synaptic targeting of AMPA-type glutamate (GluR1) receptors, and decreased AMPA receptor-mediated synaptic transmission. Reduction of endogenous neurabin levels by interference RNA (RNAi)-mediated knockdown also inhibited the surface expression of GluR1 receptors. Together, our studies suggested that disrupting the functions of a cytoskeletal neurabin/PP1 complex enhanced filopodia and impaired surface GluR1 expression in hippocampal neurons, thereby hindering the morphological and functional maturation of dendritic spines.
Figures
Similar articles
-
Targeting protein phosphatase 1 (PP1) to the actin cytoskeleton: the neurabin I/PP1 complex regulates cell morphology.Mol Cell Biol. 2002 Jul;22(13):4690-701. doi: 10.1128/MCB.22.13.4690-4701.2002. Mol Cell Biol. 2002. PMID: 12052877 Free PMC article.
-
Actin-associated neurabin-protein phosphatase-1 complex regulates hippocampal plasticity.J Neurochem. 2006 Sep;98(6):1841-51. doi: 10.1111/j.1471-4159.2006.04070.x. Epub 2006 Aug 8. J Neurochem. 2006. PMID: 16899074
-
Differential regulation of AMPA receptor trafficking by neurabin-targeted synaptic protein phosphatase-1 in synaptic transmission and long-term depression in hippocampus.J Neurosci. 2007 Apr 25;27(17):4674-86. doi: 10.1523/JNEUROSCI.5365-06.2007. J Neurosci. 2007. PMID: 17460080 Free PMC article.
-
Regulation of actin dynamics in dendritic spines: Nanostructure, molecular mobility, and signaling mechanisms.Mol Cell Neurosci. 2020 Dec;109:103564. doi: 10.1016/j.mcn.2020.103564. Epub 2020 Oct 20. Mol Cell Neurosci. 2020. PMID: 33096206 Review.
-
New waves in dendritic spine actin cytoskeleton: From branches and bundles to rings, from actin binding proteins to post-translational modifications.Mol Cell Neurosci. 2017 Oct;84:77-84. doi: 10.1016/j.mcn.2017.05.002. Epub 2017 May 4. Mol Cell Neurosci. 2017. PMID: 28479292 Review.
Cited by
-
Regulation of Synaptic Transmission and Plasticity by Protein Phosphatase 1.J Neurosci. 2021 Apr 7;41(14):3040-3050. doi: 10.1523/JNEUROSCI.2026-20.2021. J Neurosci. 2021. PMID: 33827970 Free PMC article. Review.
-
Actin and Actin-Binding Proteins: Masters of Dendritic Spine Formation, Morphology, and Function.Open Neurosci J. 2009 Jan 1;3:54-66. doi: 10.2174/1874082000903020054. Open Neurosci J. 2009. PMID: 20717495 Free PMC article.
-
Subsynaptic AMPA receptor distribution is acutely regulated by actin-driven reorganization of the postsynaptic density.J Neurosci. 2012 Jan 11;32(2):658-73. doi: 10.1523/JNEUROSCI.2927-11.2012. J Neurosci. 2012. PMID: 22238102 Free PMC article.
-
cAMP regulation of protein phosphatases PP1 and PP2A in brain.Biochim Biophys Acta Mol Cell Res. 2019 Jan;1866(1):64-73. doi: 10.1016/j.bbamcr.2018.09.006. Epub 2018 Sep 18. Biochim Biophys Acta Mol Cell Res. 2019. PMID: 30401536 Free PMC article. Review.
-
Expression and function of calcium-activated potassium channels in human glioma cells.Glia. 2006 Aug 15;54(3):223-33. doi: 10.1002/glia.20364. Glia. 2006. PMID: 16817201 Free PMC article.
References
-
- Barnes, A. P., Smith, F. D., 3rd, VanDongen, H. M., VanDongen, A. M., and Milgram, S. L. (2004). The identification of a second actin-binding region in spinophilin/neurabin II. Brain Res. Mol. Brain Res. 124, 105–113. - PubMed
-
- Beattie, E. C., Carroll, R. C., Yu, X., Morishita, W., Yasuda, H., von Zastrow, M., and Malenka, R. C. (2000). Regulation of AMPA receptor endocytosis by a signaling mechanism shared with LTD. Nat. Neurosci. 3, 1291–1300. - PubMed
-
- Bonhoeffer, T., and Yuste, R. (2002). Spine motility. Phenomenology, mechanisms, and function. Neuron 35, 1019–1027. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
