

doi: 10.1534/genetics.104.036343.
Epub 2005 Mar 2.
Synthetic lethality of retinoblastoma mutant cells in the Drosophila eye by mutation of a novel peptidyl prolyl isomerase gene
Affiliations
- PMID: 15744054
- PMCID: PMC1449713
- DOI: 10.1534/genetics.104.036343
Item in Clipboard
Synthetic lethality of retinoblastoma mutant cells in the Drosophila eye by mutation of a novel peptidyl prolyl isomerase gene
Genetics.
2005 May.
Abstract
Mutations that inactivate the retinoblastoma (Rb) pathway are common in human tumors. Such mutations promote tumor growth by deregulating the G1 cell cycle checkpoint. However, uncontrolled cell cycle progression can also produce new liabilities for cell survival. To uncover such liabilities in Rb mutant cells, we performed a clonal screen in the Drosophila eye to identify second-site mutations that eliminate Rbf(-) cells, but allow Rbf(+) cells to survive. Here we report the identification of a mutation in a novel highly conserved peptidyl prolyl isomerase (PPIase) that selectively eliminates Rbf(-) cells from the Drosophila eye.
Figures
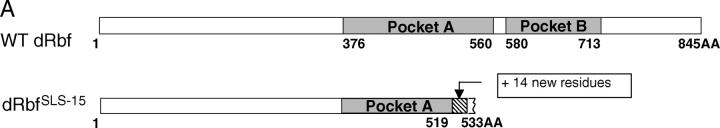
Schematics of the Rbf protein and Rbf rescue construct. (A) Diagram of the wild-type Rbf and RbfSLS-15 mutant proteins. The mutation analysis of the RbfSLS-15 transcripts revealed an 11-bp deletion resulting in a frameshift at amino acid residue 519, followed by the addition of 14 novel residues and truncation of the Rbf protein at residue 533. The truncated protein lacks Pocket B, a highly conserved RBF domain that is required for interactions with partner proteins and the execution of RBF function. (B) Diagram of the Rbf rescue construct and Rbf− clone generation. The RbfSLS-15 mutation combined with a Rbf rescue construct allows for the generation of Rbf− clones specifically in the eye, due to eye-specific FLP expression followed by recombination between the FRT sites and subsequent loss of the Rbf+ and w+ genes. All other tissues, which do not express FLP, remain Rbf+, resulting in a rescue of the organismal lethality normally associated with Rbf-deficient flies.

Schematics of the Rbf protein and Rbf rescue construct. (A) Diagram of the wild-type Rbf and RbfSLS-15 mutant proteins. The mutation analysis of the RbfSLS-15 transcripts revealed an 11-bp deletion resulting in a frameshift at amino acid residue 519, followed by the addition of 14 novel residues and truncation of the Rbf protein at residue 533. The truncated protein lacks Pocket B, a highly conserved RBF domain that is required for interactions with partner proteins and the execution of RBF function. (B) Diagram of the Rbf rescue construct and Rbf− clone generation. The RbfSLS-15 mutation combined with a Rbf rescue construct allows for the generation of Rbf− clones specifically in the eye, due to eye-specific FLP expression followed by recombination between the FRT sites and subsequent loss of the Rbf+ and w+ genes. All other tissues, which do not express FLP, remain Rbf+, resulting in a rescue of the organismal lethality normally associated with Rbf-deficient flies.
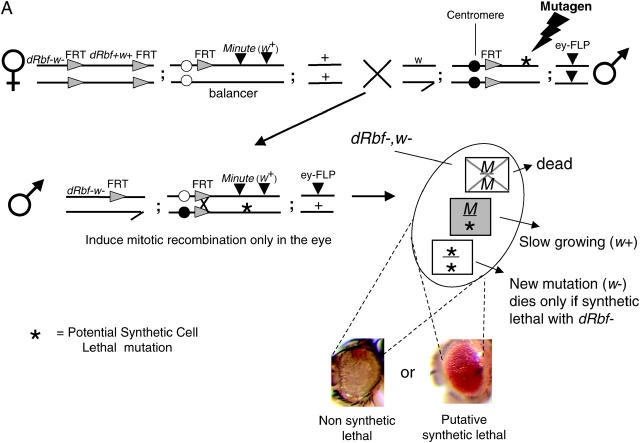
Schematic of the primary screen and counterscreen. (A) Schematic of the primary screen. Rbf+ screening-stock virgin females were crossed to mutagenized male stocks. Male progeny were assayed for mutations that resulted in the loss of w eye clones, causing the eyes to be w+. Two separate FLP/FRT recombination events are initiated by the eyeless promoter. First, the FRTs flanking the Rbf rescue construct recombine in cis, eliminating the Rbf+ and w+ genes, resulting in a large Rbf−, w clone in the eye. Second, the trans recombination between the two autosomal FRTs results in the generation of three different cell types: 1. Minute/Minute (M/M): This cell type is cell lethal because M/M cells die, regardless of the Rbf status of the cell. 2. Minute/mutation (M/*): This cell type is viable and marked with w+. When cells are heterozygous for Minute they are slow growing and are easily outcompeted. 3. mutation/mutation (*/*): This cell type is viable if the mutation is not synthetic lethal with Rbf−, since this outcompetes the M/* clone, resulting in a 90–95% w eye. When there is a synthetic lethal interaction with Rbf−, the clone is unable to populate the eye and M/* is the only cell type that survives, resulting in a w+ eye. (B) Schematic of the counterscreen. To eliminate those mutations that are not dependent upon Rbf status, hits from the primary screen were crossed to Rbf+ MFRT line virgins. The FRT/FLP recombination events under the direction of the eyeless promoter result in the generation of three different cell types: (1) M/M, as described above; (2) M/*, as described above; and (3) */*, if the previously observed synthetic lethal phenotype is indeed Rbf− specific, this cell type will be able to populate the eye in a Rbf+ background, resulting in a w eye. Conversely, if these cells are absent, resulting in a w+ eye, then there is no Rbf− synthetic interaction and the previously observed phenotype was due to nonspecific cell lethality.
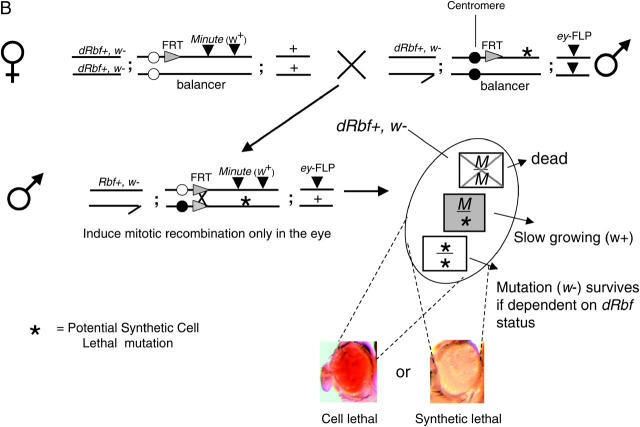
Schematic of the primary screen and counterscreen. (A) Schematic of the primary screen. Rbf+ screening-stock virgin females were crossed to mutagenized male stocks. Male progeny were assayed for mutations that resulted in the loss of w eye clones, causing the eyes to be w+. Two separate FLP/FRT recombination events are initiated by the eyeless promoter. First, the FRTs flanking the Rbf rescue construct recombine in cis, eliminating the Rbf+ and w+ genes, resulting in a large Rbf−, w clone in the eye. Second, the trans recombination between the two autosomal FRTs results in the generation of three different cell types: 1. Minute/Minute (M/M): This cell type is cell lethal because M/M cells die, regardless of the Rbf status of the cell. 2. Minute/mutation (M/*): This cell type is viable and marked with w+. When cells are heterozygous for Minute they are slow growing and are easily outcompeted. 3. mutation/mutation (*/*): This cell type is viable if the mutation is not synthetic lethal with Rbf−, since this outcompetes the M/* clone, resulting in a 90–95% w eye. When there is a synthetic lethal interaction with Rbf−, the clone is unable to populate the eye and M/* is the only cell type that survives, resulting in a w+ eye. (B) Schematic of the counterscreen. To eliminate those mutations that are not dependent upon Rbf status, hits from the primary screen were crossed to Rbf+ MFRT line virgins. The FRT/FLP recombination events under the direction of the eyeless promoter result in the generation of three different cell types: (1) M/M, as described above; (2) M/*, as described above; and (3) */*, if the previously observed synthetic lethal phenotype is indeed Rbf− specific, this cell type will be able to populate the eye in a Rbf+ background, resulting in a w eye. Conversely, if these cells are absent, resulting in a w+ eye, then there is no Rbf− synthetic interaction and the previously observed phenotype was due to nonspecific cell lethality.
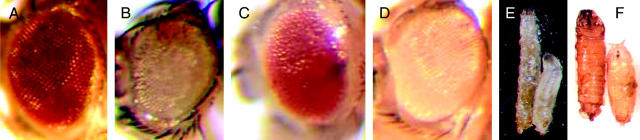
Phenotypes of KE1-1 eye clones, mutant larvae, and pupae. (A) Wild-type Drosophila eye. (B) Rbf−, w clone generated in the screening stock. (C) Clone of KE1-1 generated in the Rbf− w screening stock. The KE1-1, Rbf−, w cells die due to synthetic lethality, leaving the eye populated with Rbf−, M, w+ cells. (D) Clone of KE1-1 generated in the Rbf+ counterscreen stock. The KE1-1, Rbf+, w cells are viable, demonstrating that KE1-1 is not cell lethal on its own. (E) Large larva phenotype of a KE1-1/KE1-1 wandering third instar larva (left) compared to a KE1-1/+ larva (right). (F) Rare KE1-1/KE1-1 escaper pupae (left) are also large compared to KE1-1/+ pupae (right). Full genotypes of flies shown in B–D are: (B) RbfSLS-15, PExp{FRT2.1[Rbf+, w+, 3.5ey-FLP]}; P{ry[+t7.2] = neoFRT}42D P{w[+mC] = piM}45F M(2)53[1]/P{ry[+t7.2] = neoFRT}42D iso2; P{ry[+7.2] = ey-FLP.N}6, ry[506}]; (C) w, RbfSLS-15, Pexp{FRT2.1 [Rbf+, w+, 3.5ey-FLP]}; P{ry[+t7.2] = neoFRT}42D, P{w[+mC] = piM}45F, M(2)53[1]/ P{ry[+t7.2] = neoFRT}42D, iso2[KE1-1]; P{ry[+7.2] = ey-FLP.N}6, ry[506]/+; (D) w; P{ry[+t7.2] = neoFRT}42D, P{w[+mC] = piM}45F, M(2)53[1]/P{ry[+t7.2] = neoFRT}42D, iso2[KE1-1]; P{ry[+7.2] = ey-FLP.N}6, ry[506]/+.
CG3511 encodes a unique and highly conserved peptidyl prolyl isomerase protein. (A) The KE1-1 mutant contains a two-nucleotide substitution and a single-base-pair deletion in the transcript of CG3511-RA, when compared to wild type. A partial sequence of the transcript between nucleotides 545 and 600 is shown, with the changes present in the KE1-1 mutant given in boldface type. (B) Protein sequence alignment of CG3511 and its predicted human ortholog KIAA0073. Identical residues are shaded in black, similar residues are shaded gray. The WD domains and prolyl isomerase domain predictions are graphically represented above the alignment by hatched bars and solid bars, respectively. An asterisk denotes the location of the first frameshifted residue in the KE1-1 mutant. (C) Conservation of predicted proteins and domains encoded by the KE1-1 allele, wild-type CG3511, and selected eukaryotic orthologs. PPIL1 represents the next closest PPIase to CG3511 and is shown for comparison. The organization of WD motifs and the peptidyl prolyl isomerase within the proteins is depicted by boxes. Percentage sequence identities throughout the proteins and within the conserved peptidyl prolyl isomerase domains are shown.

CG3511 encodes a unique and highly conserved peptidyl prolyl isomerase protein. (A) The KE1-1 mutant contains a two-nucleotide substitution and a single-base-pair deletion in the transcript of CG3511-RA, when compared to wild type. A partial sequence of the transcript between nucleotides 545 and 600 is shown, with the changes present in the KE1-1 mutant given in boldface type. (B) Protein sequence alignment of CG3511 and its predicted human ortholog KIAA0073. Identical residues are shaded in black, similar residues are shaded gray. The WD domains and prolyl isomerase domain predictions are graphically represented above the alignment by hatched bars and solid bars, respectively. An asterisk denotes the location of the first frameshifted residue in the KE1-1 mutant. (C) Conservation of predicted proteins and domains encoded by the KE1-1 allele, wild-type CG3511, and selected eukaryotic orthologs. PPIL1 represents the next closest PPIase to CG3511 and is shown for comparison. The organization of WD motifs and the peptidyl prolyl isomerase within the proteins is depicted by boxes. Percentage sequence identities throughout the proteins and within the conserved peptidyl prolyl isomerase domains are shown.
CG3511 encodes a unique and highly conserved peptidyl prolyl isomerase protein. (A) The KE1-1 mutant contains a two-nucleotide substitution and a single-base-pair deletion in the transcript of CG3511-RA, when compared to wild type. A partial sequence of the transcript between nucleotides 545 and 600 is shown, with the changes present in the KE1-1 mutant given in boldface type. (B) Protein sequence alignment of CG3511 and its predicted human ortholog KIAA0073. Identical residues are shaded in black, similar residues are shaded gray. The WD domains and prolyl isomerase domain predictions are graphically represented above the alignment by hatched bars and solid bars, respectively. An asterisk denotes the location of the first frameshifted residue in the KE1-1 mutant. (C) Conservation of predicted proteins and domains encoded by the KE1-1 allele, wild-type CG3511, and selected eukaryotic orthologs. PPIL1 represents the next closest PPIase to CG3511 and is shown for comparison. The organization of WD motifs and the peptidyl prolyl isomerase within the proteins is depicted by boxes. Percentage sequence identities throughout the proteins and within the conserved peptidyl prolyl isomerase domains are shown.
F2 lethal noncomplementation screen for additional KE1 alleles. Mutagenized yw; FRT(42D); ey-FLP males were mated to females bearing additional copies of ey-FLP. Single male F1 progeny, heterozygous for the newly induced mutations, were mated to KE1-1 females and the F2 progeny were scored for the absence of [FRT(42D)*/KE1-1] flies.
CG3511 is underexpressed in KE1 mutants. Quantitative analysis of CG3511 transcript levels in larvae is shown. The y-axis shows normalized CG3511 transcript levels (see materials and methods ) present in wild-type (IsoFSR), KE1-1, and KE1-2 mutant third instar larvae. The reduction in transcript levels observed in the KE1-2 larvae is >10-fold.
References
-
- Andreotti, A. H., 2003. Native state proline isomerization: an intrinsic molecular switch. Biochemistry 42: 9515–9524. - PubMed
-
- Chau, B. N., and J. Y. Wang, 2003. Coordinated regulation of life and death by RB. Nat. Rev. Cancer 3: 130–138. - PubMed
-
- Cryns, V. L., A. Thor, H. J. Xu, S. X. Hu, M. E. Wierman et al., 1994. Loss of the retinoblastoma tumor-suppressor gene in parathyroid carcinoma. N. Engl. J. Med. 330: 757–761. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
