

Coexistence of bacterial sulfide oxidizers, sulfate reducers, and spirochetes in a gutless worm (Oligochaeta) from the Peru margin
- PMID: 15746360
- PMCID: PMC1065125
- DOI: 10.1128/AEM.71.3.1553-1561.2005
Coexistence of bacterial sulfide oxidizers, sulfate reducers, and spirochetes in a gutless worm (Oligochaeta) from the Peru margin
Abstract
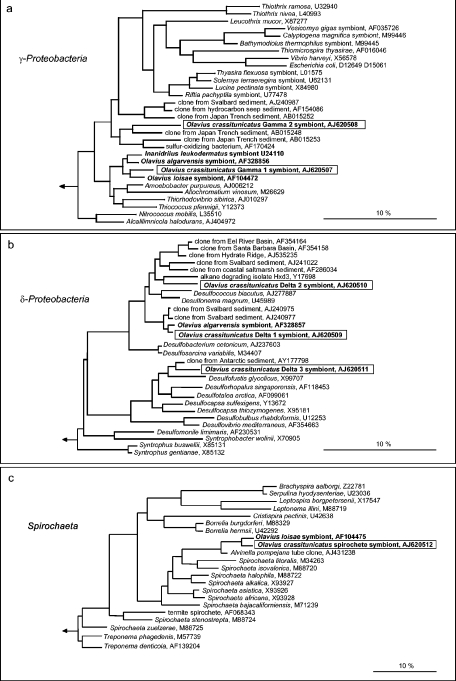
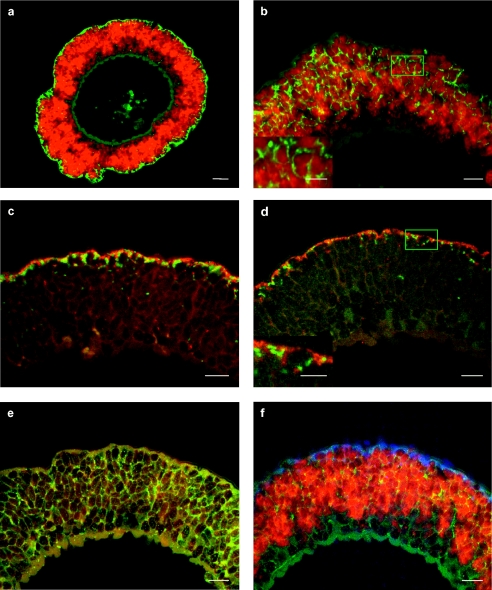
Olavius crassitunicatus is a small symbiont-bearing worm that occurs at high abundance in oxygen-deficient sediments in the East Pacific Ocean. Using comparative 16S rRNA sequence analysis and fluorescence in situ hybridization, we examined the diversity and phylogeny of bacterial symbionts in two geographically distant O. crassitunicatus populations (separated by 385 km) on the Peru margin (water depth, approximately 300 m). Five distinct bacterial phylotypes co-occurred in all specimens from both sites: two members of the gamma-Proteobacteria (Gamma 1 and 2 symbionts), two members of the delta-Proteobacteria (Delta 1 and 2 symbionts), and one spirochete. A sixth phylotype belonging to the delta-Proteobacteria (Delta 3 symbiont) was found in only one of the two host populations. Three of the O. crassitunicatus bacterial phylotypes are closely related to symbionts of other gutless oligochaete species; the Gamma 1 phylotype is closely related to sulfide-oxidizing symbionts of Olavius algarvensis, Olavius loisae, and Inanidrilus leukodermatus, the Delta 1 phylotype is closely related to sulfate-reducing symbionts of O. algarvensis, and the spirochete is closely related to spirochetal symbionts of O. loisae. In contrast, the Gamma 2 phylotype and the Delta 2 and 3 phylotypes belong to novel lineages that are not related to other bacterial symbionts. Such a phylogenetically diverse yet highly specific and stable association in which multiple bacterial phylotypes coexist within a single host has not been described previously for marine invertebrates.
Figures
References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Breznak, J. A. 2002. Phylogenetic diversity and physiology of termite gut spirochetes. Intergr. Comp. Biol. 42:313-318. - PubMed
-
- de Bary, A. 1879. Die Erscheinung der Symbiose. Verlag von Karl J. Trubner, Strassburg, Germany.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
