

Comparative analysis of selected genes from Diachasmimorpha longicaudata entomopoxvirus and other poxviruses
- PMID: 15749105
- PMCID: PMC7094658
- DOI: 10.1016/j.jinsphys.2004.10.010
Comparative analysis of selected genes from Diachasmimorpha longicaudata entomopoxvirus and other poxviruses
Abstract
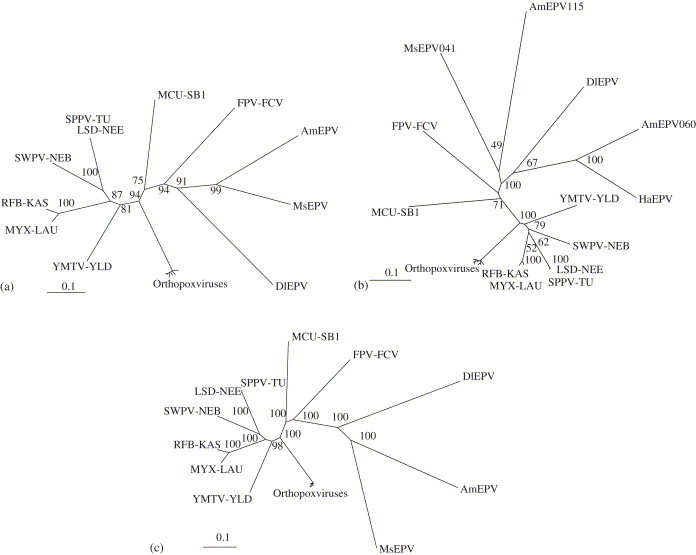
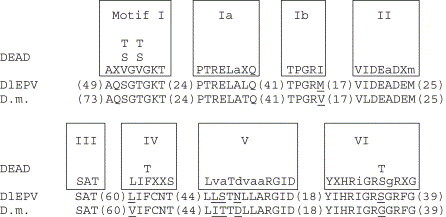
The Diachasmimorpha longicaudata entomopoxvirus (DlEPV) is the first symbiotic EPV described from a parasitic wasp. The DlEPV is introduced into the tephritid fruit fly larval host along with the wasp egg at oviposition. We sequenced a shotgun genomic library of the DlEPV DNA and analyzed and compared the predicted protein sequences of eight ORFs with those of selected poxviruses and other organisms. BlastP searches showed that five of these are homologous to poxvirus putative proteins such as metalloprotease, a putative membrane protein, late transcription factor-3, virion surface protein, and poly (A) polymerase (PAP) regulatory small subunit. Three of these are similar to those of other organisms such as the gamma-glutamyltransferase (GGT) of Arabidopsis thaliana, eukaryotic initiation factor 4A (eIF4A) of Caenorhabditis briggsae and lambda phage integrase (lambda-Int) of Enterococcus faecium. Transcription motifs for early (TGA,A/T,XXXXA) or late (TAAATG, TAAT, or TAAAT) gene expression conserved in poxviruses were identified with those ORFs. Phylogenetic analysis of multiple alignments of five ORFs and 20 poxvirus homologous sequences and of a concatenate of multiple alignments suggested that DlEPV probably diverged from the ancestral node between the fowlpox virus and the genus B, lepidopteran and orthopteran EPVs, to which Amsacta moorei and Melanoplus sanguinipes EPV, respectively, belong. The DlEPV putative GGT, eIF4A, and lambda-Int contained many conserved domains that typified these proteins. These homologues may be involved in either viral pathogenicity or enhancing parasitism via the gamma-glutamyl cycle and compensation of eIF4A levels in the parasitized fly, or via the integration of a portion of the viral genome into the wasp and/or parasitized fly.
Figures



Similar articles
-
A homolog of the vaccinia virus D13L rifampicin resistance gene is in the entomopoxvirus of the parasitic wasp, Diachasmimorpha longicaudata.J Insect Sci. 2008;8:8. doi: 10.1673/031.008.0801. J Insect Sci. 2008. PMID: 20345294 Free PMC article.
-
Purification and partial characterization of an entomopoxvirus (DLEPV) from a parasitic wasp of tephritid fruit flies.J Insect Sci. 2002;2:10. doi: 10.1093/jis/2.1.10. Epub 2002 May 28. J Insect Sci. 2002. PMID: 15455044 Free PMC article.
-
A putative DNA helicase and novel oligoribonuclease in the Diachasmimorpha longicaudata entomopoxvirus (DlEPV).Arch Virol. 2003 Jul;148(7):1431-44. doi: 10.1007/s00705-002-0975-3. Arch Virol. 2003. PMID: 12827471
-
A Mutualistic Poxvirus Exhibits Convergent Evolution with Other Heritable Viruses in Parasitoid Wasps.J Virol. 2020 Mar 31;94(8):e02059-19. doi: 10.1128/JVI.02059-19. Print 2020 Mar 31. J Virol. 2020. PMID: 32024779 Free PMC article.
-
Origin and evolution of polydnaviruses by symbiogenesis of insect DNA viruses in endoparasitic wasps.J Insect Physiol. 2003 May;49(5):419-32. doi: 10.1016/s0022-1910(03)00059-3. J Insect Physiol. 2003. PMID: 12770621 Review.
Cited by
-
Review of Venoms of Non-Polydnavirus Carrying Ichneumonoid Wasps.Biology (Basel). 2021 Jan 12;10(1):50. doi: 10.3390/biology10010050. Biology (Basel). 2021. PMID: 33445639 Free PMC article. Review.
-
Glutathione levels modulation as a strategy in host-parasite interactions-insights for biology of cancer.Front Pharmacol. 2014 Aug 5;5:180. doi: 10.3389/fphar.2014.00180. eCollection 2014. Front Pharmacol. 2014. PMID: 25140152 Free PMC article. No abstract available.
-
Genomic analysis reveals an exogenous viral symbiont with dual functionality in parasitoid wasps and their hosts.PLoS Pathog. 2020 Nov 30;16(11):e1009069. doi: 10.1371/journal.ppat.1009069. eCollection 2020 Nov. PLoS Pathog. 2020. PMID: 33253317 Free PMC article.
-
An Amsacta moorei entomopoxvirus ortholog of the poly(A) polymerase small subunit exhibits methyltransferase activity and is non-essential for virus growth.Virology. 2008 Jun 5;375(2):624-36. doi: 10.1016/j.virol.2008.02.023. Epub 2008 Mar 26. Virology. 2008. PMID: 18367228 Free PMC article.
-
A homolog of the vaccinia virus D13L rifampicin resistance gene is in the entomopoxvirus of the parasitic wasp, Diachasmimorpha longicaudata.J Insect Sci. 2008;8:8. doi: 10.1673/031.008.0801. J Insect Sci. 2008. PMID: 20345294 Free PMC article.
References
-
- Abremski K.E., Hoess R.H. Evidence for a second conserved arginine residue in the integrase family of recombination proteins. Protein Engineering. 1992;5:87–91. - PubMed
Further reading
-
- Lawrence, P.O., 2000. Parasitism specific proteins and a symbiotic entomopoxvirus from Diachasmimorpha longicaudata, a parasite of tephritid fruit flies. Abstracts of the XXI International Congress of Entomology and XVIII Brazilian Congress of Entomology, Iguassu Falls, Brazil. Abstract #2371, Book II, p. 600.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
