

Molecular genetic evidence for the role of SGT1 in the intramolecular complementation of Bs2 protein activity in Nicotiana benthamiana
- PMID: 15749757
- PMCID: PMC1088001
- DOI: 10.1105/tpc.104.029637
Molecular genetic evidence for the role of SGT1 in the intramolecular complementation of Bs2 protein activity in Nicotiana benthamiana
Abstract
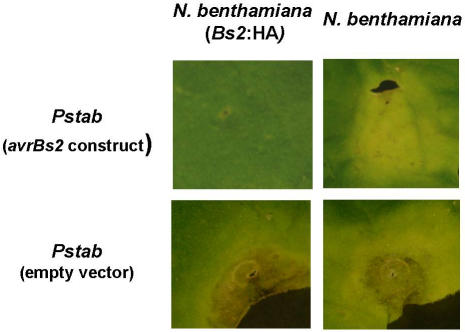
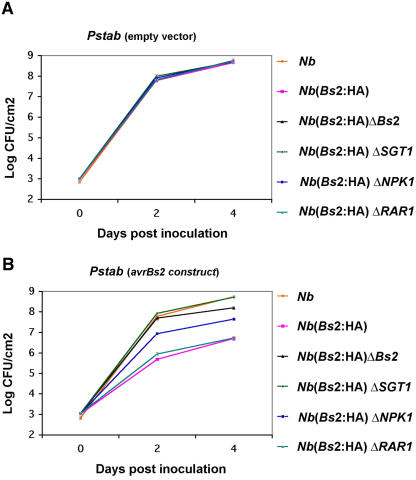
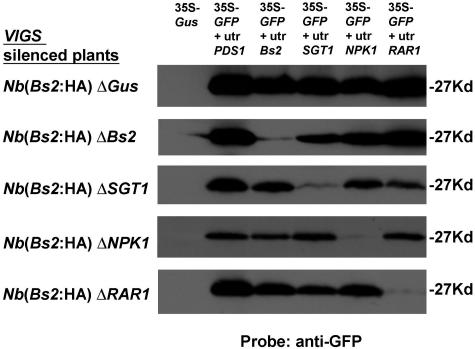
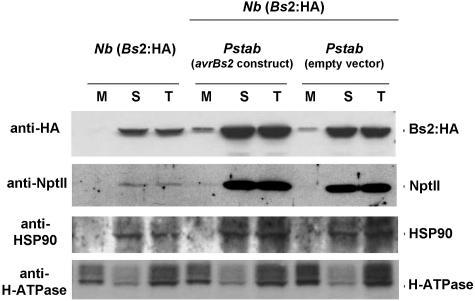
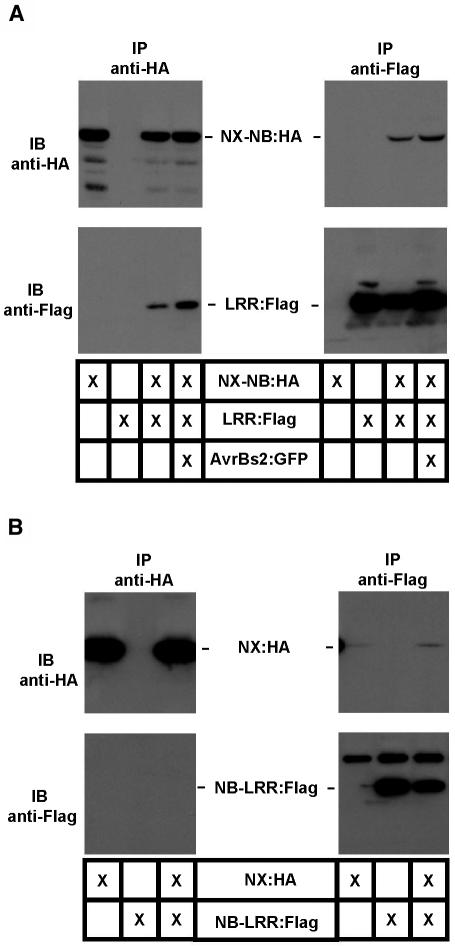
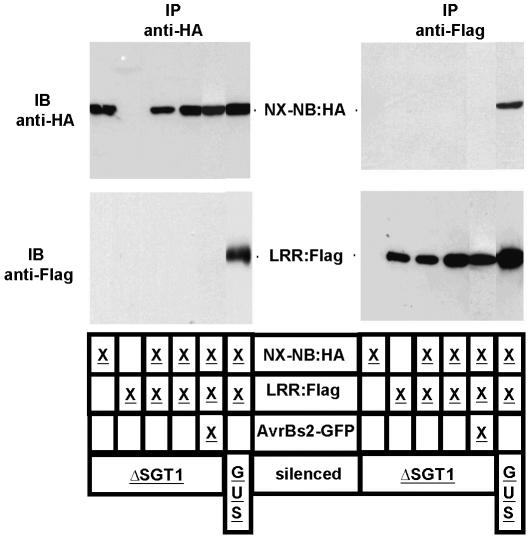
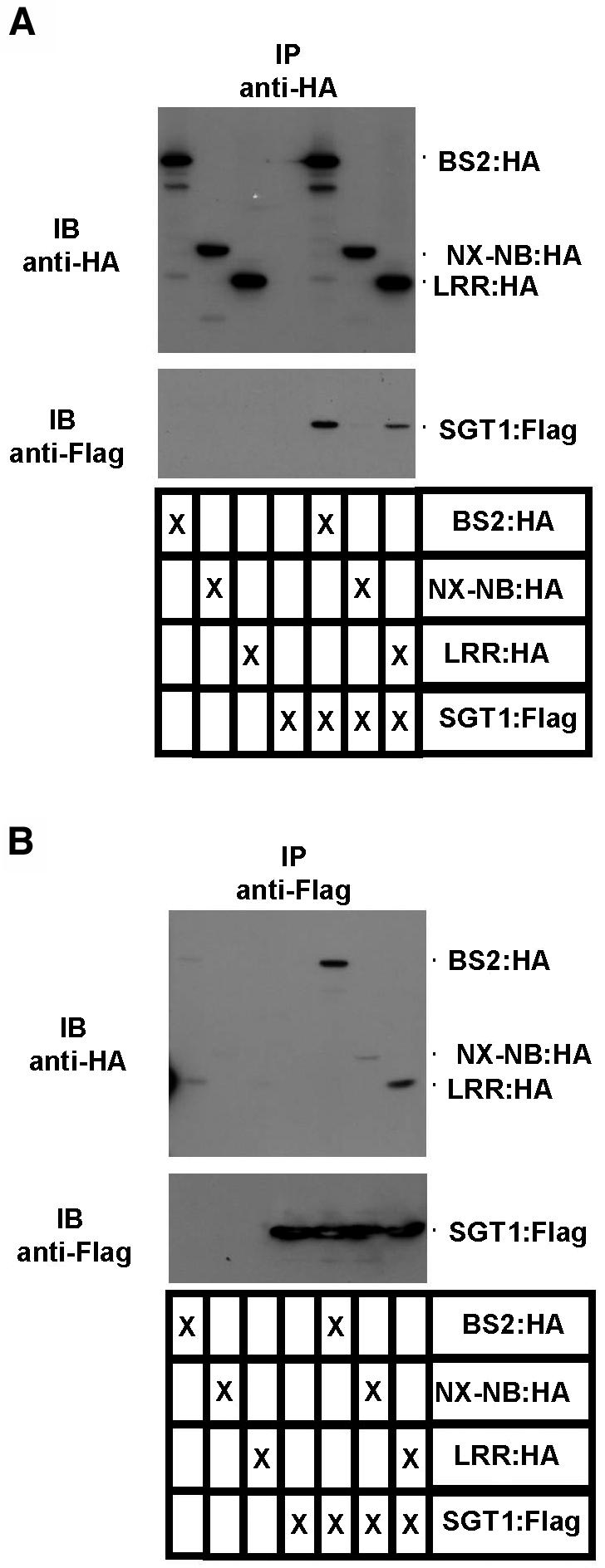
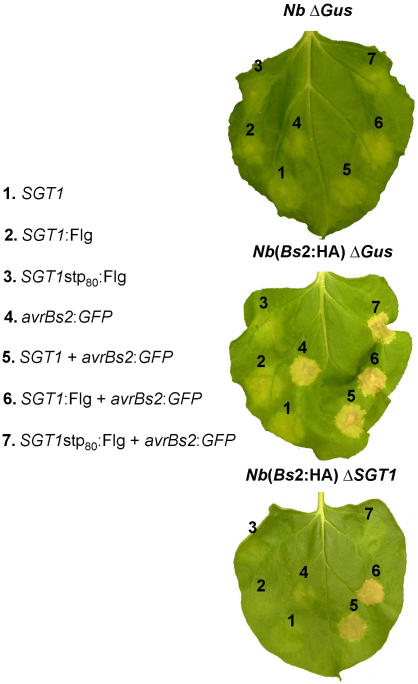
Pepper plants (Capsicum annuum) containing the Bs2 resistance gene are resistant to strains of Xanthomonas campestris pv vesicatoria (Xcv) expressing the bacterial effector protein AvrBs2. AvrBs2 is delivered directly to the plant cell via the type III protein secretion system (TTSS) of Xcv. Upon recognition of AvrBs2 by plants expressing the Bs2 gene, a signal transduction cascade is activated leading to a bacterial disease resistance response. Here, we describe a novel pathosystem that consists of epitope-tagged Bs2-expressing transgenic Nicotiana benthamiana plants and engineered strains of Pseudomonas syringae pv tabaci that deliver the effector domain of the Xcv AvrBs2 protein via the TTSS of P. syringae. This pathosystem has allowed us to exploit N. benthamiana as a model host plant to use Agrobacterium tumefaciens-mediated transient protein expression in conjunction with virus-induced gene silencing to validate genes and to identify protein interactions required for the expression of plant host resistance. In this study, we demonstrate that two genes, NbSGT1 and NbNPK1, are required for the Bs2/AvrBs2-mediated resistance responses but that NbRAR1 is not. Protein localization studies in these plants indicate that full-length Bs2 is primarily localized in the plant cytoplasm. Three protein domains of Bs2 have been identified: the N terminus, a central nucleotide binding site, and a C-terminal Leu-rich repeat (LRR). Co-immunoprecipitation studies demonstrate that separate epitope-tagged Bs2 domain constructs interact in trans specifically in the plant cell. Co-immunoprecipitation studies also demonstrate that an NbSGT1-dependent intramolecular interaction is required for Bs2 function. Additionally, Bs2 has been shown to associate with SGT1 via the LRR domain of Bs2. These data suggest a role for SGT1 in the proper folding of Bs2 or the formation of a Bs2-SGT1-containing protein complex that is required for the expression of bacterial disease resistance.
Figures
References
-
- Axtell, M.J., and Staskawicz, B.J. (2003). Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112, 369–377. - PubMed
-
- Azevedo, C., Sadanandom, A., Kitagawa, K., Freialdenhoven, A., Shirasu, K., and Schulze-Lefert, P. (2002). The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance. Science 295, 2073–2076. - PubMed
-
- Bieri, S., Mauch, S., Shen, Q.H., Peart, J., Devoto, A., Casais, C., Ceron, F., Schulze, S., Steinbiss, H.-H., Shirasu, K., and Schulze-Lefert, P. (2004). RAR1 positively controls steady state levels of barley MLA resistance proteins and enables sufficient MLA6 accumulation for effective resistance. Plant Cell 16, 3480–3495. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
